
Isolation of Bacillus amyloliquefaciens JK6 and identification of its lipopeptides surfactin for suppressing tomato bacterial wilt†
Hanqin
Xiong
,
Yongtao
Li
,
Yanfei
Cai
*,
Yu
Cao
and
Yan
Wang
College of Natural Resources and Environment, South China Agricultural University, Guangzhou, 510642, PR China. E-mail: yanfeicai@scau.edu.cn; xionghanqinxyz@163.com; Tel: +86 13535219941
First published on 21st September 2015
Abstract
A rhizobacteria strain, B. amyloliquefaciens JK6, isolated from the rhizosphere soil of healthy tomato plants, significantly inhibited Ralstonia solanacearum (RS). The biocontrol efficacies (BCEs) of JK6 suppressing tomato bacterial wilt caused by RS were up to 58.6% and 52.9% in two greenhouse experiments. To evaluate the mechanisms of the antagonist, we found that JK6 could produce siderophores, protease, biofilm, cellulose, indole acetic acid (IAA) and ammonium (NH3). In addition, PCR was used to identify antimicrobial genes in JK6. Amplification products of the expected sizes were identified as srfAB, involved in surfactin synthesis; ituA, ituB, ituC and ituD, involved in iturin synthesis; fenD, involved in fengycin synthesis; and yndJ, involved in the biosynthesis of the Yndj protein. The concentration of the lipopeptides surfactin isolated from JK6 culture was 64.24 mg L−1 identified by liquid chromatography coupled with mass spectroscopy. Furthermore, the srfAB, fenD and yndJ DNA content (copy number per 100 ng of total soil DNA) in the JK6-treated soil was significantly higher than that of the controls, as determined through real-time PCR analysis. In summary, we estimated that the production of lipopeptides surfactin as well as the higher srfAB, fenD and yndJ DNA copy numbers detected in the rhizosphere soil may play major roles in the biocontrol mechanisms with which JK6 protects plants from pathogen attacking.
1. Introduction
Ralstonia solanacearum (RS) is one of the most devastating soil-borne bacterial plant pathogens, with a worldwide distribution and an extensive host range of more than 450 plant species in 54 families.1 The RS can seriously threaten tomato growth and cause huge losses in most countries throughout the world. There are no effective chemicals to control it. Although chemical fungicides are usually used as a solution, their repeated and abusive use in the past several decades have led to a lot of environmental and food quality problems. Therefore, biocontrol represents an attractive alternative for plant disease management because of their environment-friendly way of working. Bacillus species, particularly B. subtilis and B. amyloliquefaciens, are the most effective biological control agents (BCAs) to control soil-borne plant pathogens through their various mechanisms.2 They can produce a wide range of antimicrobial compounds like lipopeptides antibiotic, including iturin, surfactin and fengycin,3 siderophores, biofilm,4 lytic enzymes such as catalase, protease and cellulase.5As biocontrol mechanism for BCAs, the product of lipopeptides antibiotic plays an important role in the biocontrol activity. The variety of lipopeptides antibiotic that some Bacillus strains naturally synthesized may explain why they are more efficient upon inhibiting plant diseases than others. Cao et al. have identified that the lipopeptides of fengycin and bacillomycin produced by B. subtilis SQR9 were effectively suppressing Fusarium wilt of cucumber.6 Cawoy et al. have also found a strong correlation between defense-inducing activity and the amount of surfactin produced by the isolates.7 Surfactin was shown to be required for biofilm development8 and may also synergistically impact the anti-fungal activity of other lipopeptides antibiotic, such as fengycin and iturin.9 Therefore, lipopeptides antibiotic production could be an efficient approach to select good candidates as biopesticides. However, what should the effect be when surfactin is produced by B. amyloliquefaciens to suppress RS?
The genome of B. amyloliquefaciens, e.g., FZB42 and AS43.3, harbours an array of giant gene clusters involved in the synthesis of lipopeptides antibiotic and other antimicrobial compounds.10,11B. amyloliquefaciens FZB42 dedicates approximately 8.5% of its total genetic capacity to the synthesis of antimicrobial compounds.12 The genes clusters, srf, fenD, bmy, were shown to direct the synthesis of the cyclic lipopeptides surfactin, fengycin, and bacillomycin. Cao et al. screened B. subtilis SQR9's genome for identifying genes involved in biosynthesis of antibiotics.6 Abdulwareth studied the quantitative expression of four lipopeptides antibiotic biosynthesis genes (ituC, srfAA, fenD and bacA) in Bacillus strains during their in vitro interaction with RS.13 The results indicated that various biosynthesis genes in the Bacillus strains Am1 and D16 exhibited different expression levels in their co-culture with RS compared with those observed in mono-culture. These results may correlate with the potential antagonistic mechanisms of these genes. Ling et al. monitored the DNA copy number of plant pathogens and biocontrol bacteria in the rhizosphere soil to evaluate the dynamic change of populations, and came up with some working mechanisms of what occurred between the microbes and plant roots in soil.14 However, few studies, if any, have been devoted to investigating antimicrobial genes expression in the rhizosphere soil in their interactions with plants.15 So it is novel and important for us to explore the potential relationship between the colonization quantity of antimicrobial genes and biocontrol efficacies in the rhizosphere soil.
In view of the above, the aims of this study are (1) to isolate and identify the novel broad-spectrum antagonistic bacteria B. amyloliquefaciens JK6 from the rhizosphere soil for healthy tomato plants; (2) to evaluate the biocontrol efficacy and biomass promotion of JK6 against RS under greenhouse conditions; (3) to isolate and identify the antibacterial compounds produced by JK6 through biochemical and molecular biology methodologies to elucidate the biocontrol mechanisms of JK6 against RS; and (4) to detect the DNA amount of antimicrobial genes in the rhizosphere soil during their interaction with plants through real-time PCR analysis, which may improve our understanding of the biocontrol mechanisms of JK6 against RS.
2. Materials and methods
2.1. Isolation and identification of antagonistic strain
Antagonistic strains were isolated from the rhizosphere soil of healthy tomato plants in Guangdong Province, China. The soil sample (10 g) was shaken in 90 mL of sterilized water for 30 min and then serially diluted and spread on Luria-Bertani medium (LB).16 After 48 h of incubation at 30 °C, single bacterial colonies were selected and streaked onto a new LB plate. The purified colonies were preserved in LB agar containing 15% glycerol at −80 °C for further studies.The antagonistic activity of the isolates suppressing RS was evaluated by the dual inoculation technique described by Lam et al. with a slight modification.17 Holes (5 mm) were punched in the middle of the test plates, which were prepared with LB agar medium mixed with RS (106 cfu mL−1), and each of the potential bacterial suspensions (40 μL, 107 cfu mL−1) was added to the holes in triplicate. The antagonistic activity was determined by the widths of the clear zone after incubation at 30 °C for 48 h.
The identification of molecular approach of strain was performed by evaluating the 16S rRNA gene sequence. Amplification was carried out by PCR using two universal primers, namely 27F and 1492R.18 The comparison of the sequence similarity was performed using Blastn, and some related species of the 16S rRNA sequence were downloaded and aligned using Mega6.0. The unrooted tree was constructed using the neighbour-joining method with a bootstrap value of 1000 replicates.
2.2. Inhibition spectrum of JK6
Strain JK6 was also evaluated for its in vitro potential to inhibit several other soil-borne pathogens, including F. oxysporum f.sp.cubense, Colletotrichum gloeosporioides, Peronophythora litchii, Magnaporthe oryzae and F. oxysporum f.sp.cucumerinum. The fungal pathogenic strains were incubated on PDA plates at 28 °C for five days. A 5 mm-diameter block of mycelium was placed in the centre of a new PDA plate. Strain JK6 was spotted with toothpicks at a distance 2.5 cm away from the rim of the mycelia colony and cultivated at 28 °C for five days. The antagonistic activities were evaluated by subtracting the distance of the fungal growth in the direction of JK6 strain from the fungal growth radius of a control culture without JK6. The experiments were repeated three times and each test had three replicates.2.3. Product of antibacterial metabolites by strain JK6
Siderophores was evaluated with the method described by Shin with a slight modification. Solutions A and B were prepared as described by Shin et al. and autoclaved at 121 °C for 20 min. The two solutions were mixed before being added to the plates. The JK6 strain was cultivated at 30 °C for two days, and siderophores product was evaluated by a colour change in the agar from blue to orange.19 Protease activity was detected using casein degradation, which can be evaluated by a distinct zone around the colony after incubating for two days at 30 °C in skim milk agar plates.5In vitro biofilm development assays were performed as described previously.20 Cellulase activity was determined by the method described by Yang.5 The IAA assay of JK6 was performed using the method described by Bric.212.4. Pot experiment design
The BCEs of JK6 for suppressing tomato bacterial wilt were determined under greenhouse conditions. The treatments were as follows: control, only inoculated with RS; JK6 treatment, inoculated with RS and JK6. The tomato seedlings were transplanted into pots until they had five leaves. In seven days after transplanting, the soil used in the JK6 treatment was inoculated with the JK6 bacterial suspension, which was resuspended with sterile water to obtain a final concentration of 106 cells per g of soil. Two days later, all the plant roots were artificially wounded and inoculated with 15 mL of RS suspension (108 cfu mL−1) to reach a final concentration of 106 cells per g of soil. The disease incidences (DIs) and plant growth promotion were measured on the 30th day after transplanting. Each treatment had 24 replicates that were in a completely randomized design and were watered regularly. The entire experiment was performed twice in an Artificial Climate Chamber (PQX-450R-22HM) with temperatures of 25–32 °C and a relative humidity from 65% to 80%. Spring pot experiment was carried out from March to May and autumn pot experiment was carried out from September to November in 2014.The typical symptoms of bacterial wilt were evaluated daily with a disease index (di) on a scale of 0 to 4, as described by Tans-Kersten.22 The DI was calculated according to Guo as follows:23 DI = [∑(number of diseased plants in the index × di)/(total number of plants investigated × highest di)] × 100%. The biocontrol efficacy was calculated as follows: biocontrol efficacy = [(DI of control − DI of antagonist treated group)]/DI of control × 100%.
2.5. Amplification of antimicrobial genes from JK6
The primers (Table S1†) used for the amplification of the JK6 antimicrobial genes were described previously by Joshi and Gardener24 and Cao.6 Each PCR reaction consisted of 2 μL of template DNA, 19 μL of sterile distilled water, 25 μL Premix (rTaq) (TAKARA) and 2 μL of each primer. PCR was performed with a cycle of 94 °C for 5 min followed by 30 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 45 s, and an extension step at 72 °C for 10 min was conducted after 30 amplification cycles. Each PCR reaction had a negative control without DNA and three replicates. The amplified products were visualized by gel electrophoresison a 1.2% agarose gel stained with ethidium bromide. The expected fragments were sequenced by Invitrogen Inc. (Shanghai). Analysis of the sequenced nucleotides was performed with the Blastn software in GenBank.2.6. Soil collection and real-time PCR analysis
Rhizosphere soil samples located 0–1.5 mm from the rhizoplane were collected on the 23rd day after inoculating with JK6 from each treatment. The total soil DNA was isolated by use of the MoBio PowerSoil® DNA Isolation Kit (MoBio Laboratories, Carlsbad, CA, USA).The abundant antimicrobial genes yndJ, fenD, ituC and srfAB in the soil DNA were determined by real-time PCR analysis. The ituC gene level was quantified by use of the primer pair ITUC-F and ITUC-R.13 The primer pairs used for yndJ, fenD and srfAB gene quantification are described in Table S1.† The real-time PCR amplification was performed by use of Thermal Cycler Dice TP800 (TaKaRa) and Bestar™ SYBR Green qPCR (DBI) master mix. The real-time conditions were selected according to the manufacturer's protocol with appropriate modifications. The procedure started with an initial denaturation step at 95 °C for 2 min, and this step was followed by 40 cycles of 95 °C for 15 s and 60 °C for 20 s. The primer specificity and the formation of primer dimers were monitored by melt curve analysis. All of the experiments were performed in triplicates. The plasmid standards used for antimicrobial gene quantification were generated from the target genes cloned from JK6 genomic DNA. Standard curves were generated according to the previous report.25 The abundant yndJ, fenD, ituC and srfAB genes were expressed with their copy concentration.
2.7. Identification and quantification of lipopeptides surfactin from JK6
Antimicrobial compounds were extracted from a 48 h-old culture of JK6 as described previously by Dunlap with a slight modification. Briefly, the cell-free culture filtrate was partitioned into fractions by use of a C18 syringe cartridge (Bond Elut C18, Agilent, USA) and then washed with 10 mL of water, and fractions were eluted with 1 mL of 100% methanol.11 The C18-fractionated supernatant mixtures were prepared by passing the pale-yellow solution. The antibiotics were detected by a mass of spectrographic instruments (Agilent 6410 Triple Quadrupole LC/MS, Agilent, USA) for molecular weight determination. The MS analysis was performed by electrospray ionization in the negative ion mode.26According to the results of the MS analysis, we evaluated that JK6 may harbour the lipopeptides surfactin. The level of lipopeptides surfactin was quantified by use of the method described by Wang with reversed-phase HPLC analysis.27 Commercial surfactin (Sigma-Aldrich St. Louis, USA) was used as a standard.
2.8. Statistical analyses
All data obtained were statistically analysed with the SPSS 20.0 program. The data were subjected to Duncan's analysis of variance (ANOVA), and the means were separated by Duncan's multiple range tests at P < 0.05.3. Results
3.1. Isolation and identification of antagonistic bacterial strains
In total, 58 rhizobacteria were isolated from the soil samples, and eight of these showed inhibitory effects against RS, ranging from 4 to 34.7 mm. JK6 exerted the highest inhibitory effect on RS in vitro. Furthermore, we found that JK6 significantly suppressed the growth of F. oxysporum f.sp.cubense, Magnaporthe oryzae, Colletotrichum gloeosporioides, Peronophythora litchii and F. oxysporum f.sp.cucumerinum on PDA plates (Fig. 1). The antifungal spectra of JK6 ranged from 34.3 to 43.3 mm, as shown in Table 1. Therefore, the JK6 strain was used in the subsequent studies.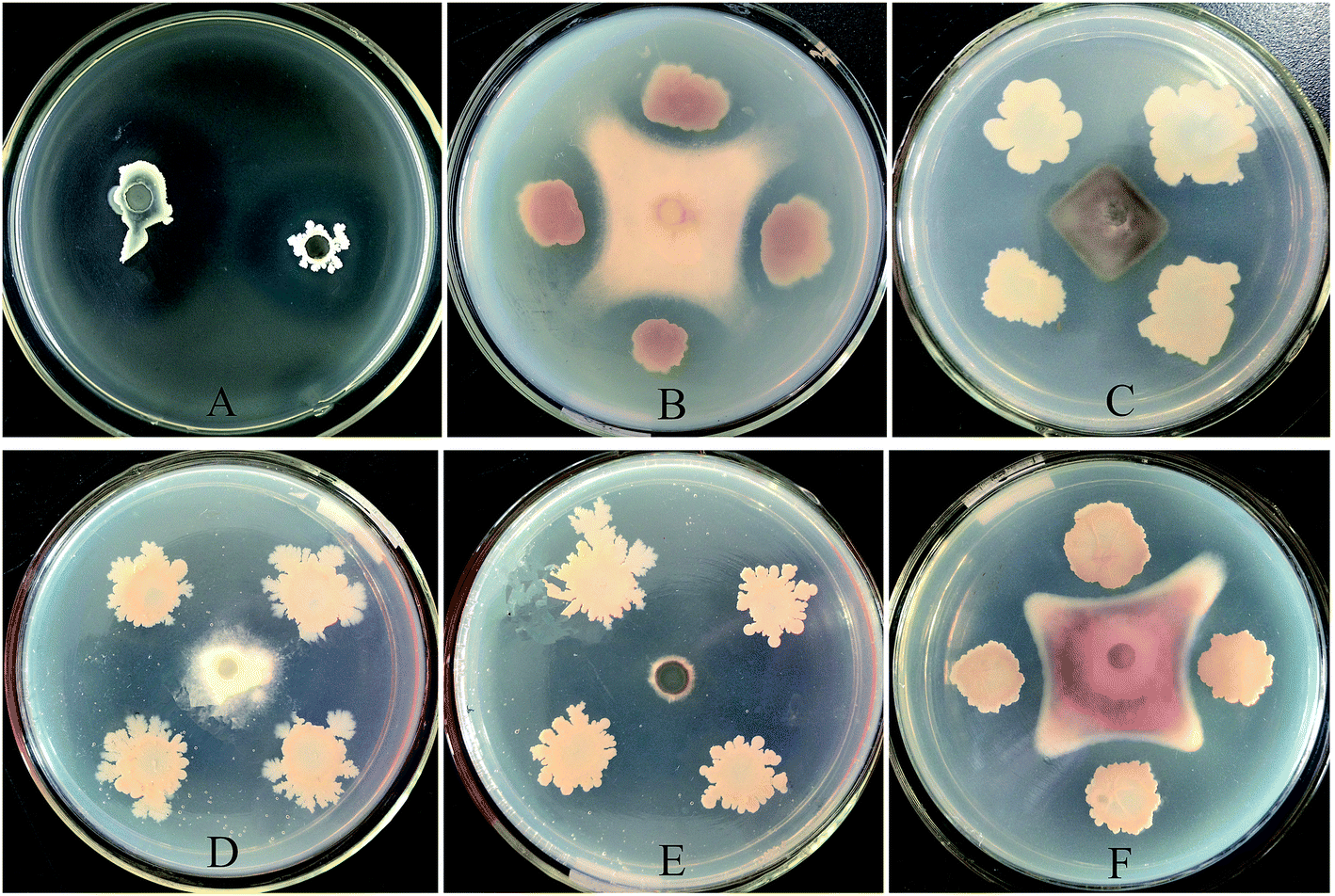 | ||
| Fig. 1 Demonstration of the inhibitory effects of B. amyloliquefaciens JK6 against RS (A), F. oxysporum f.sp.cubense (B), Colletotrichum gloeosporioides (C), Peronophythora litchii (D), Magnaporthe oryzae (E) and F. oxysporum f.sp. cucumerinum (F). For the antagonism with RS, bacterial suspension (40 μL, 107 cfu mL−1) of JK6 was added into the holes in the test plates, which were prepared with LB agar medium mixed with RS (106 cfu mL−1). Plates were incubated at 30 °C for 48 h to observe the inhibition zones. The fungal pathogenic strains were incubated on PDA plates at 28 °C for 5–7 days until sporulation. A 5 mm block of mycelium was cut and transferred into a new PDA plate. The JK6 strain was spotted with toothpicks 2.5 cm away from the fungal block and cultivated at 28 °C for 5 days. | ||
| Plant pathogens | Inhibition diameter (mm) |
|---|---|
| a The suppression diameters are the mean values ± SE for three replicates. The data in the column with different capital letters differ significantly at P < 0.05 by Duncan's test. | |
| R. solanacearum | 34.7 ± 0.3 c |
| F. oxysporum f.sp.cubense | 36.7 ± 1.3 b |
| Colletotrichum gloeosporioides | 42.0 ± 0.6 a |
| Peronophythora litchii | 34.3 ± 0.3 c |
| Magnaporthe oryzae | 43.3 ± 0.3 a |
| F. oxysporum f.sp. cucumerinum | 36.8 ± 0.2 b |
The 16S rRNA sequence of JK6 (KR 149334) was more than 99% identical to B. amyloliquefaciens FZB42, as determined by Blastn. Furthermore, the sequences of 16S rRNA in each genome were downloaded and clustered with JK6 rRNA with Mega6.0. The results clearly demonstrated that JK6 was B. amyloliquefaciens (Fig. 2).
 | ||
| Fig. 2 Phylogenetic tree of B. amyloliquefaciens strain JK6 based on 16S rRNA gene sequences and related bacteria with the neighbor-joining method with bootstrap value of 1000 replicates. The bar indicates an estimated sequence divergence of 0.2%. | ||
3.2. Evaluation of biocontrol efficacy in greenhouse condition
The DI was significantly reduced in the JK6 treatment in greenhouse condition. Specifically, the DIs in JK6 treatment were 26.7% and 41.7% in two separate experiments, which were much lower than those of the controls (64.6% and 88.5%). The BCEs of JK6 were up to 58.6% and 52.9%. Furthermore, the plant heights and dry weights in JK6 treatment were markedly higher than those of the controls, as is showed in Table 2. The results indicated that JK6 can effectively inhibit tomato bacteria wilt caused by RS and significantly promote the growth of tomato plants.| Pot trial | Treatments | Plant height (cm) | Average dry weight per plant (g) | Disease incidence (%) | Biocontrol efficacy (%) |
|---|---|---|---|---|---|
| a Control (only inoculated with RS), JK6 (treatment inoculated with RS and JK6). The dry weights were recorded after the plants were dried in an oven at 60 °C for three days at the end of the experiments. The data are the mean values ± SE. ** is significantly different (P < 0.01, Student's t-test). | |||||
| Spring | Control | 16.9 ± 0.2 | 0.54 ± 0.03 | 64.6 ± 5.8** | 58.6 |
| JK6 | 20.2 ± 0.5** | 0.77 ± 0.06** | 26.7 ± 4.3 | ||
| Autumn | Control | 23.0 ± 0.2 | 0.72 ± 0.03 | 88.5 ± 5.8** | 52.9 |
| JK6 | 25.3 ± 0.7** | 0.96 ± 0.01** | 41.7 ± 4.5 | ||
3.3. Production of antibacterial metabolites by strain JK6
The marked colour change from blue to orange on CAS agar indicated siderophores product (Fig. 3A), and the clearness of the skim milk agar suggested the strong protease activity of JK6 (Fig. 3B). The trace of crystal violet in the PVC tube demonstrated the biofilm development of JK6, showing its potential colonization ability on plant roots (Fig. 3C), and the clear zones surrounding the JK6 colony on cellulose agar plates demonstrated the cellulase activity of JK6 (Fig. 3D). JK6 also produced IAA at a rate of 3.65 μg mL−1. | ||
| Fig. 3 Production of antibacterial metabolites of JK6. Siderophores production on CAS plate (A). Protease activity on skim milk medium (B). Biofilm development in a plastic eppendorf tube (C). Cellulase activity on a cellulose agar plate (D). | ||
3.4. Identification of antimicrobial genes of JK6
A total of seven gene fragments of the anticipated size were efficiently amplified from JK6 through PCR analysis (Fig. 4). The amplicon obtained using the 147F/147R primer pair showed 95% identity with yndJ, a gene encoding the Yndj protein. The products amplified with the FNDF1/FNDR1 primer pair were similar to fengycin synthetase (98% of homology). The surfactin synthesis gene was observed in JK6 with the 110F/110R primer pair and had a homology of 96%. In addition, the PCR fragments amplified by the ituD2F/ituD2R, bamB1F/bamB1R, ituA1F/ituA1R and ITUCF1/ITUCR3 primer pairs showed 97–99% identity with regions associated with iturin synthesis (Table S2†). | ||
| Fig. 4 Agarose gel-electrophoresis of PCR fragments of antimicrobial genes from B. amyloliquefaciens JK6. The Lane 1, DL 2000 marker; Lane 2, yndJ; Lane 3, fenD; Lane 4, ituC; Lane 5, srfAB; Lane 6, ituA; Lane 7, ituD; Lane 8, ituB. | ||
3.5. Gene detection in the rhizosphere soil
The DNA quantities (copy number/100 ng total soil DNA) of the yndJ, fenD and srfAB genes in JK6 inoculated treatment with 23 days were markedly higher than those observed in the control treatment. Among these elevated genes in JK6 treatment, yndJ gene was the highest with 177![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 368 copies per 100 ng total soil DNA. The second was fenD gene with 176981 copies per 100 ng total soil DNA. SrfAB gene was the third with 934 copies. In contrast, the DNA quantities of the yndJ, fenD and srfAB genes in the control treatment were 1975, 1093 and 416 copies, respectively. However, the amount of ituC DNA showed no significant difference during these treatments (Fig. 5).
368 copies per 100 ng total soil DNA. The second was fenD gene with 176981 copies per 100 ng total soil DNA. SrfAB gene was the third with 934 copies. In contrast, the DNA quantities of the yndJ, fenD and srfAB genes in the control treatment were 1975, 1093 and 416 copies, respectively. However, the amount of ituC DNA showed no significant difference during these treatments (Fig. 5).
 | ||
| Fig. 5 DNA amounts (copy number/100 ng total soil DNA) of the yndJ, fenD, ituC and srfAB genes in the rhizosphere soil collected from the first (a) and second (b) experimental pots. Note: CK, the control treatment, only inoculated with RS; JK6, the JK6 treatment, inoculated with both RS and JK6. The DNA amounts were determined by real-time PCR analysis. The data are expressed as log10 copies per 100 ng of total soil DNA. All of the values are the means from four replicates. The bars indicate the standard deviations of the mean. * indicates a significance difference at P < 0.05, ** indicates a significance difference at P < 0.01, and *** indicates a significant difference at P < 0.001, as determined by Student's t-test. | ||
3.6. Identification and quantification of lipopeptides surfactin
The HPLC analysis showed that JK6 sample had five surfactin homologues comparing with the standard lipopeptides surfactin due to their similar peak retention time under the same HPLC conditions (Fig. 6a and b). Furthermore, the mass spectrometry analysis showed several molecules around the peaks at m/z 993.9, 1008.9, 1022.9 1036.9 and 1048.9 (Fig. 6c), suggesting a series of homologous molecules with a difference of 14-carbon tail (–CH2–), which have similar m/z values to the lipopeptides surfactin according to the literature.9 The other mass spectra, for example 992.9, 1007.9, 1021.9, 1035.9 and so on, were caused by the absence of H ions from surfactin homologous. It was easy to lose H ions because the MS analysis was performed by electrospray ionization in the negative ion mode and its structure was macrocyclic sequence (Fig. 6d).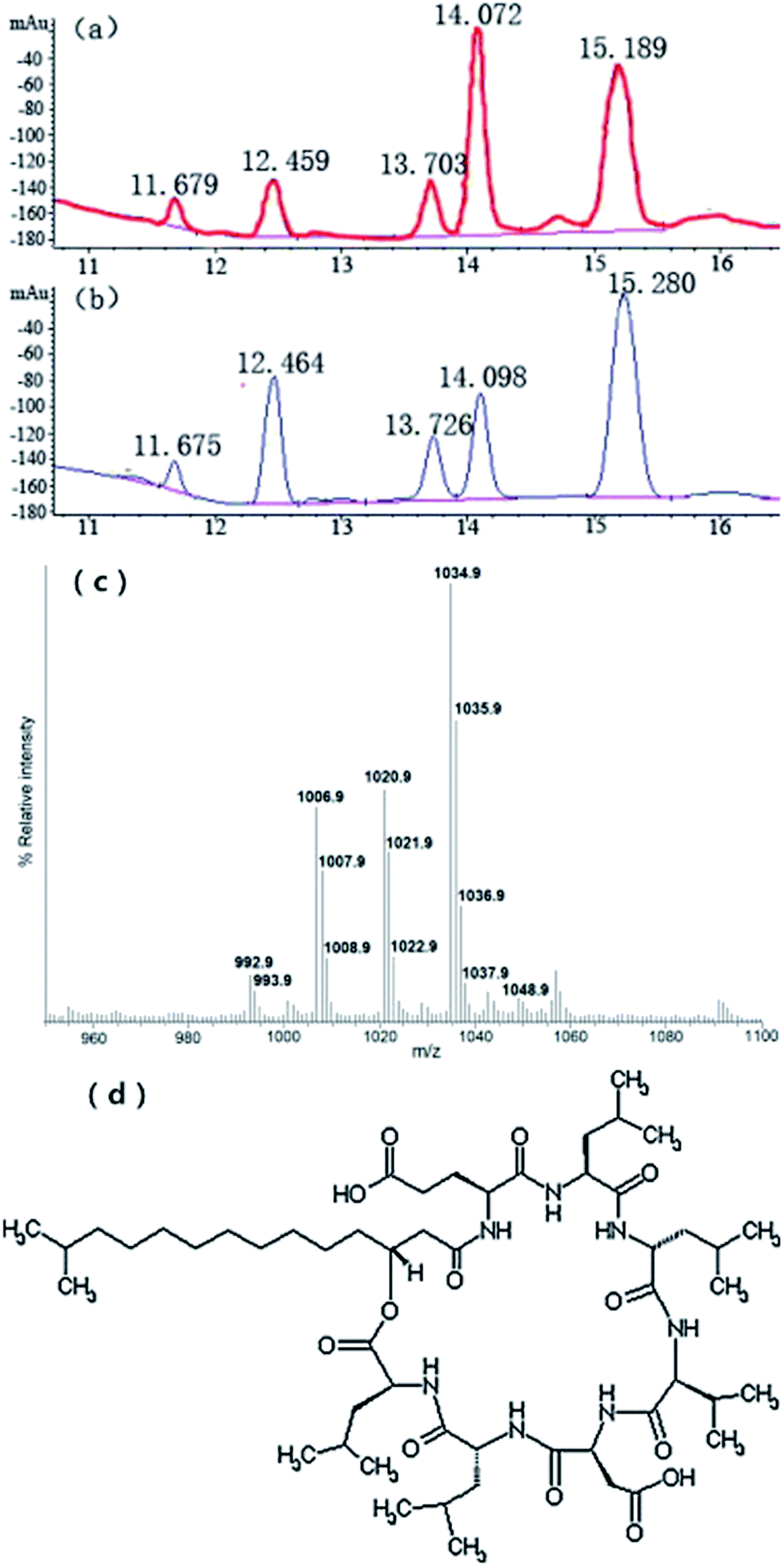 | ||
| Fig. 6 Reversed-phase HPLC chromatograms of standard lipopeptides surfactin (a) and the lipopeptides surfactin produced by B. amyloliquefaciens JK6 (b). ESI mass spectrum of lipopeptides surfactin from B. amyloliquefaciens JK6 culture in the resion m/z 950–1100 (c). Structure of lipopeptides surfactin molecule (d). | ||
The MS and HPLC analyses absolutely demonstrated that JK6 can produce lipopeptides surfactin. According to the standard curve, y = 8.0190x + 160.66 (R2 = 0.999), the lipopeptides surfactin concentration of JK6 culture was equal to 64.24 mg L−1.
4. Discussion
In this study, a novel antagonistic strain JK6 was isolated from the rhizosphere soil of healthy tomato plants and showed broad-spectrum antimicrobial activity against various plant-disease pathogens (Fig. 1). JK6 was identified as B. amyloliquefaciens through phylogenetic tree based on 16S rRNA gene sequences (Fig. 2). The inhibition diameter of JK6 against RS was up to 34.7 mm, which is markedly higher than those of B. amyloliquefaciens CM-2 (11.7 mm) and Pseudomonas J12 (24.7 mm).4,28 Its inhibition diameters of broad-spectrum antimicrobial activity ranged from 34.3 to 43.3 mm (Table 1), which are higher than those of Bg-C31 (maximum inhibition diameter of 15.3 mm),29 indicating that it may serve as a potential BCA for controlling these diseases caused by these pathogens and other plant diseases. Additionally, JK6 significantly reduced the DI of tomato bacteria wilt and promoted plant growth in greenhouse condition (Table 2). The BCE of JK6 reached 58.6%, which is higher than that found for the J12 strain (45.5%).4 Similarly, the BCE of the APF1 strain isolated by Lemessa was 60.3%, whereas the other BCEs of other five antagonistic bacteria strains were lower than those of JK6.13The main mechanisms with which BCAs suppressing plant diseases are a broad spectrum of synthesized antibiotic and antimicrobial compounds.24 Among these antibiotic and antimicrobial compounds, lipopeptides have well-recognized potential uses in biotechnology.15 Recent investigations have demonstrated that lipopeptides can not only influence the ecological fitness of the produced strain in terms of root colonization (and thereby persistence in the rhizosphere), but also have a major role in the beneficial interaction of Bacillus species with plants by stimulating host defence mechanisms.15 In this study, PCR analysis revealed the existence of antimicrobial genes in JK6 strain, such as ituA (KR 149330), ituB (KR 149331), ituC (KR 149332), ituD (KR 149333), srfAB (KR 149335), fenD (KR 149329) and yndJ (KR 149336), which are involved in iturin, fengycin, surfactin and YndJ protein synthesis. The seven cloned sequences had high degrees of similarity to homologous sequences previously identified in other biocontrol strains, i.e., FZB42,30 GB03 (ref. 24) and B946 (ref. 31) (Table S2†). The isolates with these genes present an enhanced capacity to produce lipopeptides antibiotics with inhibitory activity against soil-borne diseases.21 Some research groups have proposed that lipopeptides antibiotics production play a significant role in protection against plant diseases either by hindering the pathogens directly or by generating systemic resistance in the host plants.32,33 Our data demonstrated the presence of lipopeptides surfactin in the culture filtrate of JK6 by LC-MS analysis, and the concentration of surfactin was found to equal 64.24 mg L−1. Koumoustsi reported that the lipopeptides surfactin analogues demonstrated high levels of suppressive activity against F. oxysporum f.sp.cubense.30 Our experiments also demonstrated that the surfactin produced by JK6 significantly inhibited RS and F. oxysporum f.sp.cubense. However, other lipopeptides antibiotics, including iturin and fengycin, were not detected in the JK6 culture filtrates, even though JK6 harbours genes encoding proteins associated with the synthesis of these antibiotics (fenD and ituA, ituB, ituC, and ituD). This may be due to the use of unoptimized fermentation conditions.34 The various lipopeptides antibiotics that JK6 may produce under different fermentation conditions will be evaluated in future studies.
Recent studies demonstrated the growing importance of applying PCR analysis to antagonistic bacteria which can enable a rapid identification of potential bioactive metabolites produced by prospective biocontrol organisms. But the studies of the lipopeptides genes expression in biocontrol organisms have only recently been reported. Abdulwareth et al. reported that the srfAA, ituC and bacA biosynthesis genes in Bacillus strains Am1 showed higher expression levels during the dual-culture interaction with RS compared with those in mono-culture.13 While the expression level of fenD gene reduced during dual-culture with RS than in mono-culture. This finding suggests that the antimicrobial activity of Am1 is because of the higher expression levels of these three genes (srfAA, ituC and bacA). It may also contribute to their potential ability to secrete the corresponding lipopeptides into the surrounding niches.32 Though some researchers have reported the DNA quantity or expression of microbes and plant disease pathogens in the test plants or in the rhizosphere soil.6,14,35,36 The information on lipopeptides genes quantity during plant interaction remains unclear. It may be attributed to the fact that BCAs have been studied primarily for their biocontrol efficacy against pathogens in the field or greenhouse.37 In this study, we established a protocol for the quantification of the antimicrobial genes yndJ, fenD, ituC and srfAB in the rhizosphere soil during interaction with plants, based on real-time PCR analysis. Our results indicated that the DNA copy numbers of yndJ, srfAB and fenD genes detected in JK6 treatment were much higher than those in the control treatment, which may correlate with the colonization of JK6 in the rhizosphere soil. These results will further indicate that the potential mechanism of JK6 strain for biological activity is owing to these three higher colonization genes, which may contribute to secreting corresponding lipopeptides in the rhizosphere soil environment. This is very important for us to further understand the biocontrol mechanisms from the aspect of genetic regulation mechanisms in JK6 strain.
Last but not least, several other mechanisms have been proposed to explain the inhibition of RS by B. amyloliquefaciens. For example, siderophores and lytic enzymes secreted by antagonistic strains can reduce the growth of pathogens present in the rhizosphere.38,39 Biofilm formation may contribute to root colonization and protect plants from attacking by soil-borne pathogens.4 The auxin IAA is considered the most important plant hormone, involved in the regulation of the organogenesis, cell expansion, tropic responses, differentiation and gene regulation of plants and functions as an important signaling molecule.40,41 Ammonium produced by bacteria also has been reported that it can promote the growth of plants.42 Tan et al. have showed that the production of siderophores, IAA and ammonium could contribute to the inhibition of RS and plant growth.28 Similarly, our results demonstrated that siderophores, protease, cellulose, biofilms, IAA, and ammonia produced by JK6 may contribute to its pathogen growth inhibition and plant growth promotion.
In conclusion, B. amyloliquefaciens JK6 proved to be excellent biocontrol bacteria in inhibiting RS and promoting plant growth in greenhouse. To evaluate the mechanisms of the antagonist, JK6 was found to produce lipopeptides surfactin, siderophores, protease, biofilm, cellulose, IAA and ammonium. In addition, the DNA copy numbers of yndJ, srfAB and fenD genes of JK6 treatments were significantly higher than those observed in the control treatment. This was pursued as another major mechanism underlying the biocontrol of RS by JK6. All of these features should come together to provide better efficient disease control. Thus, JK6 is a novel biocontrol strain with great potential.
Acknowledgements
This research was financially supported by the National Natural Science Foundation of China (Grant Number: 41171209, 41471214, 41171210, U1401234) and Guangzhou Science and Technology plan projects (Grant Number: 2014A020208089).References
- E. Wicker, L. Grassart, R. Coranson-Beaudu, D. Mian, C. Guilbaud and M. Fegan, Appl. Environ. Microbiol., 2007, 71, 6790–6801 CrossRef PubMed.
- A. Perez-Garcia, D. Romero and A. de Vicente, Curr. Opin. Biotechnol., 2011, 22, 187–193 CrossRef CAS PubMed.
- T. Stein, Mol. Microbiol., 2005, 56, 845–857 CrossRef CAS PubMed.
- T. T. Zhou, D. Chen, C. Y. Li, Q. Sun, L. Z. Li, F. Liu, Q. Shen and B. Shen, Microbiol. Res., 2012, 167, 388–394 CrossRef CAS PubMed.
- W. Yang, Q. Xu, H. X. Liu, Y. P. Wang, Y. M. Wang, H. T. Yang and J. H. Guo, Biol. Control, 2012, 62, 144–151 CrossRef PubMed.
- Y. Cao, Z. H. Xu, N. Ling, Y. J. Yuan, X. M. Yang, L. H. Chen, B. Shen and Q. Shen, Sci. Hortic., 2012, 135, 32–39 CrossRef CAS PubMed.
- H. Cawoy, M. Mariutto, G. Henry, C. Fisher, N. Vasiyeva, P. Thonart, J. Dommes and M. Ongena, Mol. Plant-Microbe Interact., 2014, 27, 87–100 CrossRef CAS PubMed.
- J. Hofemeister, B. Conrad, B. Adler, B. Hofemeister, J. Feesche, N. Kucheryava, G. Steinborn, P. Franke, N. Grammel and A. Zwintscher, Mol. Genet. Genomics, 2004, 272, 363–378 CrossRef CAS PubMed.
- H. Hiraoka, O. Asaka, T. Ano and M. Shoda, J. Gen. Appl. Microbiol., 1992, 38, 635–640 CrossRef CAS.
- X. H. Chen, A. Koumoutsi, R. Scholz, A. Eisenreich, K. Schneider, I. Heinemeyer, B. Morgenstern, B. Voss, W. R. Hess, O. Reva, H. Junge, B. Voigt, P. R. Jungblut, J. Vater, R. Süssmuth, H. Liesegang, A. Strittmatter, G. Gottschalk and R. Borriss, Nat. Biotechnol., 2007, 25, 1007–1014 CrossRef CAS PubMed.
- C. A. Dunlap, M. J. Bowman and D. A. Schisler, Biol. Control, 2013, 64, 166–175 CrossRef CAS PubMed.
- X. H. Chen, A. Koumoutsi, R. Scholz, K. Schneider, J. Vater, R. Süssmuth, J. Piel and R. Borriss, J. Biotechnol., 2009, 140, 27–37 CrossRef CAS PubMed.
- A. A. Abdulwareth, K. K. Ullah, N. Zarqa, B. Li, M. A. Ali, C. L. Yang and G. L. Xie, Symbiosis, 2014, 63, 59–70 CrossRef.
- N. Ling, W. W. Zhang, S. Y. Tan, Q. W. Huang and Q. Shen, Appl. Soil. Ecol., 2012, 59, 13–19 CrossRef PubMed.
- M. Ongena and P. Jacques, Trends Microbiol., 2008, 16, 115–125 CrossRef CAS PubMed.
- F. Lemessa and W. Zeller, Biol. Control, 2007, 42, 336–344 CrossRef PubMed.
- Y. W. Lam, H. X. Wang and T. B. Ng, Biochem. Biophys. Res. Commun., 2000, 279, 74–80 CrossRef CAS PubMed.
- A. C. Redburn and B. K. C. Patel, FEMS Microbiol. Lett., 1993, 113, 81–86 CrossRef CAS PubMed.
- S. H. Shin, Y. Lim, S. E. Lee, N. W. Yang and J. H. Rhee, J. Microbiol. Methods, 2001, 44, 89–95 CrossRef CAS.
- H. L. Wei and L. Q. Zhang, Antonie van Leeuwenhoek, 2006, 89, 267–280 CrossRef PubMed.
- J. M. Bric, R. M. Bostock and S. E. Silverstone, Appl. Environ. Microbiol., 1991, 57, 535–538 CAS.
- J. Tans-Kersten, D. Brown and C. Allen, Mol. Plant-Microbe Interact., 2004, 17, 686 CrossRef CAS PubMed.
- J. H. Guo, H. Y. Qi, Y. H. Guo, H. L. Ge, L. Y. Gong, L. X. Zhang and P. H. Sun, Biol. Control, 2004, 29, 66–72 CrossRef.
- R. Joshi and B. B. M. Gardener, Phytopathology, 2006, 96, 145–154 CrossRef CAS PubMed.
- I. S. Pantelides, S. E. Tjamos, I. A. Striglis, I. Chatzipavlidis and E. J. Paplomatas, Biol. Control, 2009, 50, 30–36 CrossRef PubMed.
- J. Wang, J. Liu, X. Wang, J. Yao and Z. Yu, Lett. Appl. Microbiol., 2004, 39, 98–102 CrossRef CAS PubMed.
- Y. Wang, Z. X. Lu, X. M. Bie and F. X. Lv, Eur. Food Res. Technol., 2010, 231, 189–196 CrossRef CAS.
- S. Y. Tan, C. L. Yang, X. L. Mei, S. Y. Shen and W. Raza, Crop Prot., 2013, 43, 134–140 CrossRef PubMed.
- H. Q. Hu, X. S. Li and H. He, Biol. Control, 2010, 54, 359–365 CrossRef PubMed.
- A. Koumoutsi, X. H. Chen, A. Henne, H. Liesegang, G. Hitzeroth, P. Franke, J. Vater and R. Borriss, J. Bacteriol., 2004, 186, 1084–1096 CrossRef CAS.
- J. Blom, C. Rueckert, B. Niu, Q. Wang and R. Borriss, J. Bacteriol., 2012, 194, 1845–1846 CrossRef CAS PubMed.
- O. Asaka, T. Ano and M. Shoda, J. Ferment. Bioeng., 1996, 81, 1–6 CrossRef CAS.
- D. Romero, A. de Vicente, R. V. Rakotoaly, S. E. Dufour, J. W. Veening, E. Arrebola, F. M. Cazorla, O. P. Kuipers, M. Paquot and A. Perez-Garcia, Mol. Plant-Microbe Interact., 2007, 20, 430–440 CrossRef CAS PubMed.
- A. Tapi, M. Chollet-Imbert, B. Scherens and P. Jacques, Appl. Microbiol. Biotechnol., 2010, 85, 1521–1531 CrossRef CAS PubMed.
- D. Gizi, I. A. Stringlis, S. E. Tjamos and E. J. Paplomatas, Biol. Control, 2011, 58, 387–392 CrossRef PubMed.
- I. S. Pantelides, S. E. Tjamos, I. A. Striglis, I. Chatzipavlidis and E. J. Paplomatas, Biol. Control, 2009, 50, 30–36 CrossRef PubMed.
- J. W. Kloepper, C. M. Ryu and S. Zhang, Phytopathology, 2004, 94, 1259–1266 CrossRef CAS PubMed.
- S. Compant, C. Clement and A. Sessitsch, Soil Biol. Biochem., 2010, 42, 669–678 CrossRef CAS PubMed.
- B. Ali, A. N. Sabri, K. Ljung and S. Hasnain, Lett. Appl. Microbiol., 2009, 48, 542–547 CrossRef CAS PubMed.
- R. J. Ryu and C. L. Patten, J. Bacteriol., 2008, 190, 7200–7208 CrossRef CAS PubMed.
- V. Kumar and K. P. Singh, Bioresour. Technol., 2001, 76, 173–175 CrossRef CAS.
- E. E. Idris, D. J. Iglesias, M. Talon and R. Borriss, Mol. Plant-Microbe Interact., 2007, 20, 619–626 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Two supporting tables. See DOI: 10.1039/c5ra13142a |
| This journal is © The Royal Society of Chemistry 2015 |