
Biochemical and histological changes in the liver and gills of Nile tilapia Oreochromis niloticus exposed to Red 195 dye
Insaf Ayadiabc,
Sandra Mariza Monteirod,
Imed Regayae,
Ana Coimbrad,
Fontainhas Fernandesd,
Maria Manuel Oliveirab,
Francisco Peixotod and
Wissem Mnif*af
aLR11-ES31 Laboratory of Biotechnology and Valorisation of Bio-Geo Resources, Higher Institute of Biotechnology of Sidi Thabet, BiotechPole of Sidi Thabet, University of Manouba, 2020, Tunisia. E-mail: w_mnif@yahoo.fr; Tel: +216 98 94 73 71
bCQVR, Chemistry Department, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal
cFaculty of Sciences of Bizerte, University of Carthage, Jarzouna, Bizerte-7021, Tunisia
dCenter for the Research and Technology of Agro-Environmental and Biological Sciences, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal
eHigher Institute of Environment Sciences and Technologies of BorjCédria, University of Carthage, Tunisia
fFaculty of Sciences and Arts in Balgarn, Bisha University, PO Box 60, Balgarn-Sabt Al Olaya 61985, Saudi Arabia
First published on 25th September 2015
Abstract
The present study investigates the biochemical and morphological responses induced in the liver and gills of Nile tilapia Oreochromis niloticus by exposure to various Red 195 dye concentrations (0.05, 0.1 and 0.2 mg L−1) for various durations (7, 14 and 21 days). The histology and antioxidant activities of catalase (CAT), glutathione reductase (GR) and glutathione S-transferase (GST) were monitored and evaluated during exposure and after recovery in clean water. The results revealed that CAT activity decreased after 7 and 21 days of exposure to 0.2 mg L−1 concentrations and increased only after 7 days of exposure to high concentrations compared to the liver and gill control samples, respectively. Both organs were, however, noted to undergo a decrease in GST activity after 7 days of exposure to low Red 195 dye concentrations. Compared to the control, the gills of the tilapia exposed to 0.05 and 0.1 mg L−1 concentrations for longer periods underwent an increase in GST activity. Similarly, GR activity was higher in the liver of tilapia exposed to this dye at the sampling days, except for the highest concentration (0.2 mg L−1) after 21 days of exposure. The GR activity in the gills decreased significantly after 7 and 21 days of exposure to 0.05 and 0.1 mg L−1 concentrations, respectively. The results of the recovery group revealed that the liver and gills displayed insignificant differences in antioxidant enzyme activities. The liver and gill tissues of the fish exposed to Red 195 showed several histopathological changes. Liver damages included an increase in cytoplasmic vacuolization, disruption of endothelial lining, metabolic zonation, necrotic cells, rupture of hepatocyte membrane, and decline of higher eosinophilia. The gills also exhibited some necrotic cells, edema, lifting of filaments and lamellar epithelium, and vascular disorders, such as extreme vasodilatation and proliferation of filament epithelium. A correlation between the biochemical and histological changes of the liver and gill tissues was established, attributing the tissue and cell damages to the accumulation of hydrogen peroxide or production of other radicals via a Fenton reaction.
1. Introduction
Increasing concerns have been expressed over the serious environmental and health effects associated with the growing amounts of toxic contaminants from domestic, industrial and agricultural activities released into the aquatic environment worldwide. Synthetic dyes represent one of the major groups of toxic compounds used in various industries, including the textile and leather processing industries.1,2 The indiscriminate discharge of synthetic dyes into aquatic systems can cause adverse effects on marine ecosystems and human life. Those toxic contaminants can accumulate in various aquatic organisms, including fish, which constitute an important component of human nutrition, thus presenting serious risks to human health. Reactive Red 195 azo dye is characterized by having one or more azo bonds (–N![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) N), bound to substitutable aromatic rings. The latter rings of these dyes are responsible for intense colour, water solubility and degradation resistance under conventional wastewater treatment,3 since some of these azo dyes and their metabolites can be mutagens or carcinogens.4
N), bound to substitutable aromatic rings. The latter rings of these dyes are responsible for intense colour, water solubility and degradation resistance under conventional wastewater treatment,3 since some of these azo dyes and their metabolites can be mutagens or carcinogens.4
Due to its low-cost and ease of use compared to natural dyes, Red 195, has become one of the most widely used dyes in various industries worldwide, including the Tunisian textile industry. The wastewaters carrying this dye present strong color and high COD values since about 10–15% of dyes are dispersed in the effluent during the dyeing process, thus posing devastating effects on the ecological balance of the recipient environment and human health.5 In fact, several studies carried out in Tunisia have demonstrated the mutagenic and genotoxic effects of various dyes, including acid violet 7, acid yellow 17, and acid orange 52.6–9 Several works have also studied the effects of different dyes on fish samples.10–12 Nevertheless, little work has been performed to investigate the effects of Red 195 dye on the biochemical and morphological responses of fish.
Fish have been widely used as models to evaluate the health of aquatic ecosystems, biomonitor environmental stress, and identify possible ways to mitigate and prevent the effects of pollution.12 Liver plays an important role in vital functions of basic metabolism13 and represents the major organ for the accumulation, biotransformation and excretion of contaminants in fish.14 The gills are the main targets of direct contamination, as they play a significant role in metal uptake, storage and eventually, transfer to the internal compartments via blood transport.15 The study of their biochemical and histological changes has provided valuable insights for the monitoring of fish exposure to contaminants in laboratory and field studies.16 The exposure to contaminants in aquatic ecosystems can enhance the intracellular formation of reactive oxygen species (ROS), which can bring oxidative stress and damage to various biological systems.17,18 Oxidative stress occurs as an outcome of imbalance between ROS generation and elimination. ROS can be detoxified by enzyme defense systems, including superoxide dismutase (SOD), catalase (CAT) and selenium-dependent glutathione peroxidase (GPx), or by non-enzymatic systems through the scavenging action of reduced glutathione (GSH) and several other molecules, such as vitamins C, A and E. Organic peroxides, on the other hand, can be detoxified by the activity of glutathione S-transferase (GST).19 Several studies showed that changes in the levels of antioxidant enzyme activities could be used as possible biomarkers in different aquatic organisms.20–22
Nile tilapia, Oreochromis niloticus, has commonly used as a target biological model for the study of xenobiotic biotransformation in toxicology studies. This popular freshwater finfish species belonging to the cichlid family has several distinctive features that support its suitability for use as a model species for the assessment of aquatic pollution. It is known by its high growth rates, distinct ability to adapt to diverse diets, great resistance to diseases and handling practices, easy reproduction in captivity and prolific rate of breeding and multiplication, and tolerance to various environmental conditions.23 In fact, tilapia has previously been used as a sentinel organism for contaminants in various toxicological studies.24 Alterations in the biotransformation enzymes of this indicator species have also been used in various biomonitoring studies to reflect and assess environmental contaminants.25
The liver and gill are the main and main sensitive target organs of xenobiotic toxicity and damage for fish.26 They also play a major role in the biotransformation of xenobiotics. The sensitivity of these tissues to pesticide-induced stress is a function of the disturbed balance between the degree of oxidative stress and the antioxidant capability.26,27 Previous studies have shown that pesticides alter enzymatic and non-enzymatic antioxidant systems and induce oxidative stress in animals.27–29 The exposure to low doses of fipronil for prolonged periods has previously been reported to induce oxidative stress in the blood serum of pregnant rats and their offsprings.30 The oxidative stress-induction potential of exposure to different sublethal concentrations of chlorpyrifos has previously been investigated in the brain, liver and gill tissues of guppy fish (Poecilia reticulate).31 In fact, morphological alterations in the liver and gill tissues of fish are useful indicators for responses to environmental stressors or toxicants. The liver is the main organ of detoxification. Due to their lipophilicity, chlorpyrifos have a high rate of gill absorption. This could be a contributing factor to the sensitivity of fish to this organophosphorous pesticide.31,32 The oxidative damages and toxicity effects induced by deltamethrin (DLM) intoxication were also investigated in the liver, kidney and gills in freshwater Nile tilapia (Oreochromis niloticus).33,34
Considering the scarcity of data on the contamination status and ecological impacts of fish exposure to Red 195 dye, the present study was undertaken to investigate and evaluate the potential biochemical changes in the activity of antioxidant enzymes and morphological responses in the liver and gill of tilapia Oreochromis niloticus exposed to different concentrations of Red 195 dye for various durations under laboratory conditions.
2. Materials and methods
2.1. Fish and experimental design
The fish samples (n = 96) used in this study were healthy adult tilapia O. niloticus of both sexes (mean total length 17.14 ± 2.36 cm; mean total weight 121.54 ± 10.17 g) obtained from the aquaculture station of the University of Trás-os-Montes and Alto Douro, Portugal. The samples were randomly distributed in 12 tanks of 100 L, with water recirculation rate of 5 L min−1, temperature range 25 ± 1 °C, and controlled photoperiod of 12D:12L for 21 days. Three groups of tilapia were exposed to different concentrations of Red 195 dye (0.05, 0.1 and 0.2 mg L−1), and the other group, serving as a control, was placed under similar conditions without the xenobiotic. Each exposure condition was performed in triplicate. Two fish samples were removed from each tank each week. The tanks were provided with supplemental aeration to maintain dissolved oxygen concentration near saturation. The fish were fed daily to visual satiation with a diet that had been tested in a previous study.35 The following water quality parameters were provided: 25 ± 1 °C of temperature; pH 7.7 ± 0.2; 6 ± 1 mg L−1 of dissolved oxygen; 376 μS cm−1 of conductivity; 0.08 ± 0.06 mg L−1 of ammonium, and 0.01 ± 0.01 mg L−1 of nitrite. The Red 195 dye concentration in the water-tanks was controlled spectrophotometrically (at λ = 520 nm) and absorbance was measured using a standard curve (0–0.2 mg L−1).After 21 days of exposure period, the remaining fish samples were transferred to Red-195-dye-free water for 30 days to study recovery responses. They were then anesthetized with 1 mL of 2-phenoxyethanol L−1 water (Sigma, Barcelona, Spain), measured, weighed, and euthanized by decapitation. Their livers and gills were removed, and one randomly chosen part of each organ was fixed for microscopy (see below). The remaining part was deeply frozen for further enzymatic assays. The tissues were crushed and homogenized with phosphate buffer KH2PO4 at pH 7.4 and centrifuged at 16![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g for 20 min at 4 °C. The supernatant was used for the subsequent enzymatic assays. All experimental procedures were performed in accordance with the Portuguese law of animal welfare (Portaria no. 1005/92) and EU Directives (2010/63/EU) on the care and use of laboratory animals.
000g for 20 min at 4 °C. The supernatant was used for the subsequent enzymatic assays. All experimental procedures were performed in accordance with the Portuguese law of animal welfare (Portaria no. 1005/92) and EU Directives (2010/63/EU) on the care and use of laboratory animals.
2.2. Analytical techniques
Catalase activity (CAT) was measured by a Clark-type oxygen electrode (Hansatech®).36 The reaction medium consisted of potassium phosphate buffer (50 mM KH2PO4 pH 7.0) and hydrogen peroxide (1 M) at a final volume of 1 mL. The medium buffer was previously submitted to a nitrogen stream to decrease dissolved oxygen. A sample volume of 50 μL (diluted 100 times) was added to the reaction medium, and the mixture was incubated at 25 °C for 2 min. Reaction was initiated by the addition of hydrogen peroxide (12 mM) at a final volume of 1 mL. After 2 min of thermostatic incubation at 25 °C and stabilization, H2O2 was added to the reaction medium, and the new slope was measured. After 30 s, the enzyme extract (diluted 100 times) was added, and the new slope measured. A control test was performed without the addition of sample to determine the autodismutation of H2O2. CAT activity was expressed as mmol H2O2 per min per mg of protein.Glutathione reductase (GR) activity was determined by a Varian-Cary® 50 spectrophotometer according to the method described by Carlberg and Mannervik (1975).37 The reaction medium consisted of potassium phosphate buffer (100 mM KH2PO4 and 0.5 mM EDTA, pH 7.4), 100 mM oxidized glutathione (GSSG) and 10 mM NADPH. GSSG was added after 2 min of sample incubation to initiate the reaction. Enzyme activity was measured at 340 nm and 25 °C by NADPH oxidation. The result was expressed as mM NADPH oxidized per min per mg of protein using the molar coefficient extinction (ε) of 6.22 × 103 mM−1 cm−1.
Glutathione S-transferase (GST) activity was determined by a Varian-Cary® 50 spectrophotometer.38 The reaction medium contained potassium phosphate buffer (100 mM KH2PO4, pH 7.0), 100 mM 1-chloro-2,4-dinitrobenzene (CDNB), 10% Triton X-100 (v/v), and 100 mM GSH at a final volume of 2 mL. Recording started after 2 minutes of incubation at 25 °C. The reaction was initiated 1 minute after the onset of recording with the addition of GSH (100 mM). The variation in absorbance was determined at 340 nm, and enzyme activity was expressed in mM CDNB conjugated per min per mg of protein (ε = 9.6 × 103 M−1 cm−1).
Protein content was determined according to the method of Bradford (1976)39 using bovine serum albumin as standard. All chemicals used in the enzymatic activity assays were of analytical purity and purchased from Sigma Chemical Co.
2.3. Histology
Two gills of each fish were randomly collected and fixed in buffered formaldehyde (Panreac, Barcelona, Spain) for 24 h, dehydrated in graded ethanol series, and embedded in paraffin wax. Sagittal sections (5 μm of thickness) were cut and mounted on glass slides. Sections were deparaffinized in xylene, hydratated in ethanol and stained with hematoxylin–eosin (HE). The liver was quickly dissected, sliced into 3 mm-thick slabs, and immersed in buffered formaldehyde for 24 h, dehydrated, and embedded in paraffin; a minimum of 5 pieces were obtained. Histological sections (5 μm of thickness) were cut and stained with H&E. The histopathological changes induced by the treatment in the gill and liver organs were photographed and analyzed by light microscopy (Nikon® Labophot).2.4. Statistical analysis
The data were analyzed by the S6.0 software (StatSoft, 2001). The mean ± standard error was calculated for each experimental group. Differences between groups were tested using one-way ANOVA, followed by a Student–Newman–Keuls multiple comparison test at a 5% level of significance.3. Results
3.1. Biochemical analysis
The changes in the activities of antioxidant enzymes in the liver and gill are shown in Fig. 1 and 2, respectively. The results revealed that Red 195 exposure at all concentrations (0.05, 0.1 and 0.2 mg L−1) and for all periods altered the normal functioning of hepatic activity tilapia by consistently decreasing CAT activities (Fig. 1A) as compared to the control. Statistically significant differences were, however, observed only for the highest concentration and for two different periods (7 and 21). Furthermore, GST activities (Fig. 1B) were noted to undergo a decrease after 7 days of exposure with all concentrations, although statistically significant values were observed only for the group exposed to the lowest concentration as compared to the control group. After 14 days of exposure, no significant differences were observed between the experimental groups, except for the one exposed to Red 195 at a concentration at a concentration of 0.1 mg L−1, which showed a slight decrease. After 21 days of exposure to a concentration of 0.2 mg L−1, however, a significant increase in GST activities was observed as compared to the control. The results also revealed that glutathione reductase activity (Fig. 1C) underwent a significant increase after 7 and 14 days of exposure. After 21 days of exposure, however, this increase was statistically significant only for the lowest concentration (0.05 mg L−1).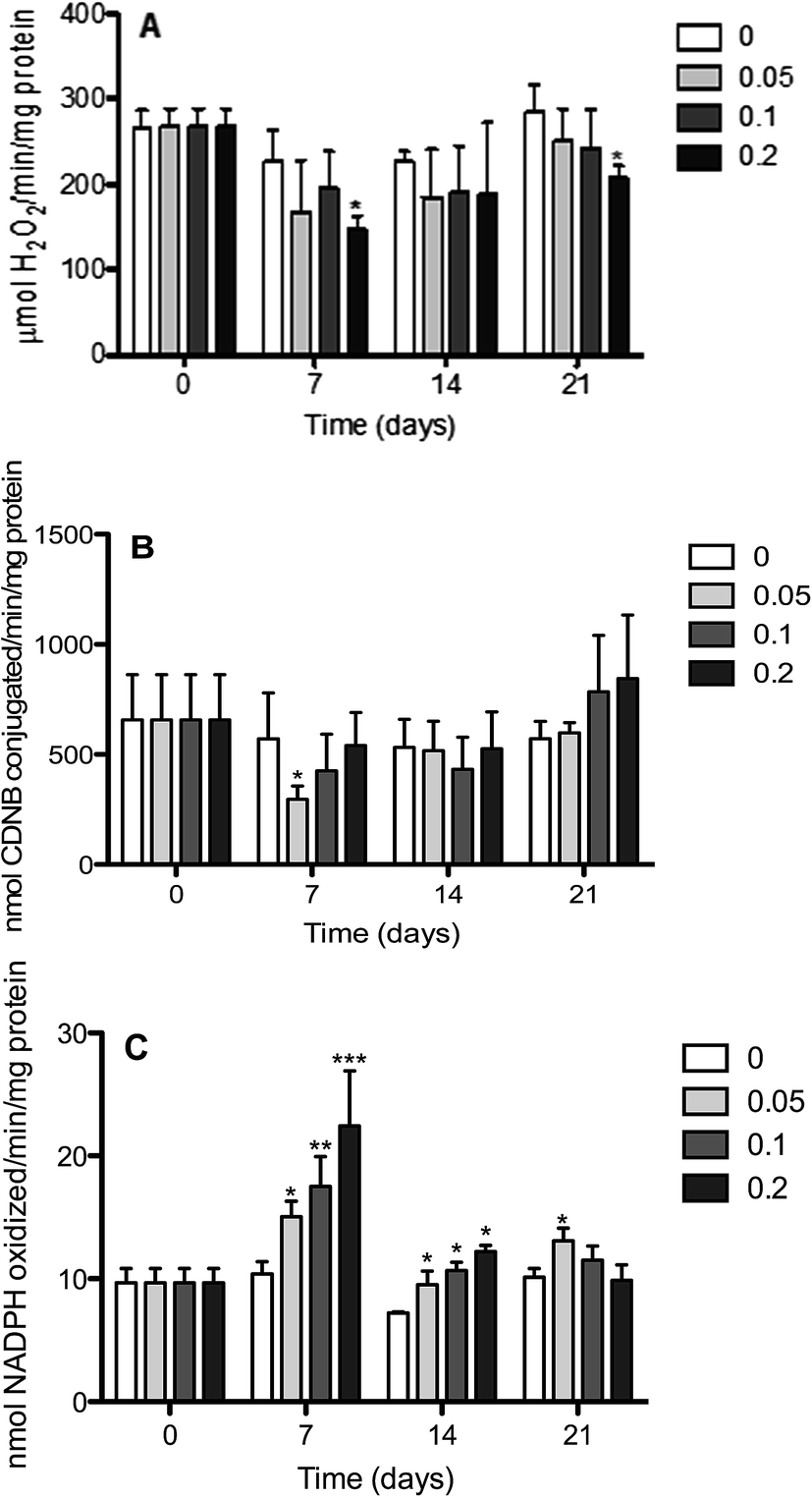 | ||
| Fig. 1 Liver antioxidant activities (means ± SD) in Nile tilapia O. niloticus exposed to different concentrations of Red 195 (0.05; 0.1 and 0.2 mg L−1). The activity levels of treated fish were compared with control group in each sampling day. Data are means ± SD of six independent experiments. *Values statistically different from control p < 0.05; **values statistically different from control p < 0.01; ***values statistically different from control p < 0.005. | ||
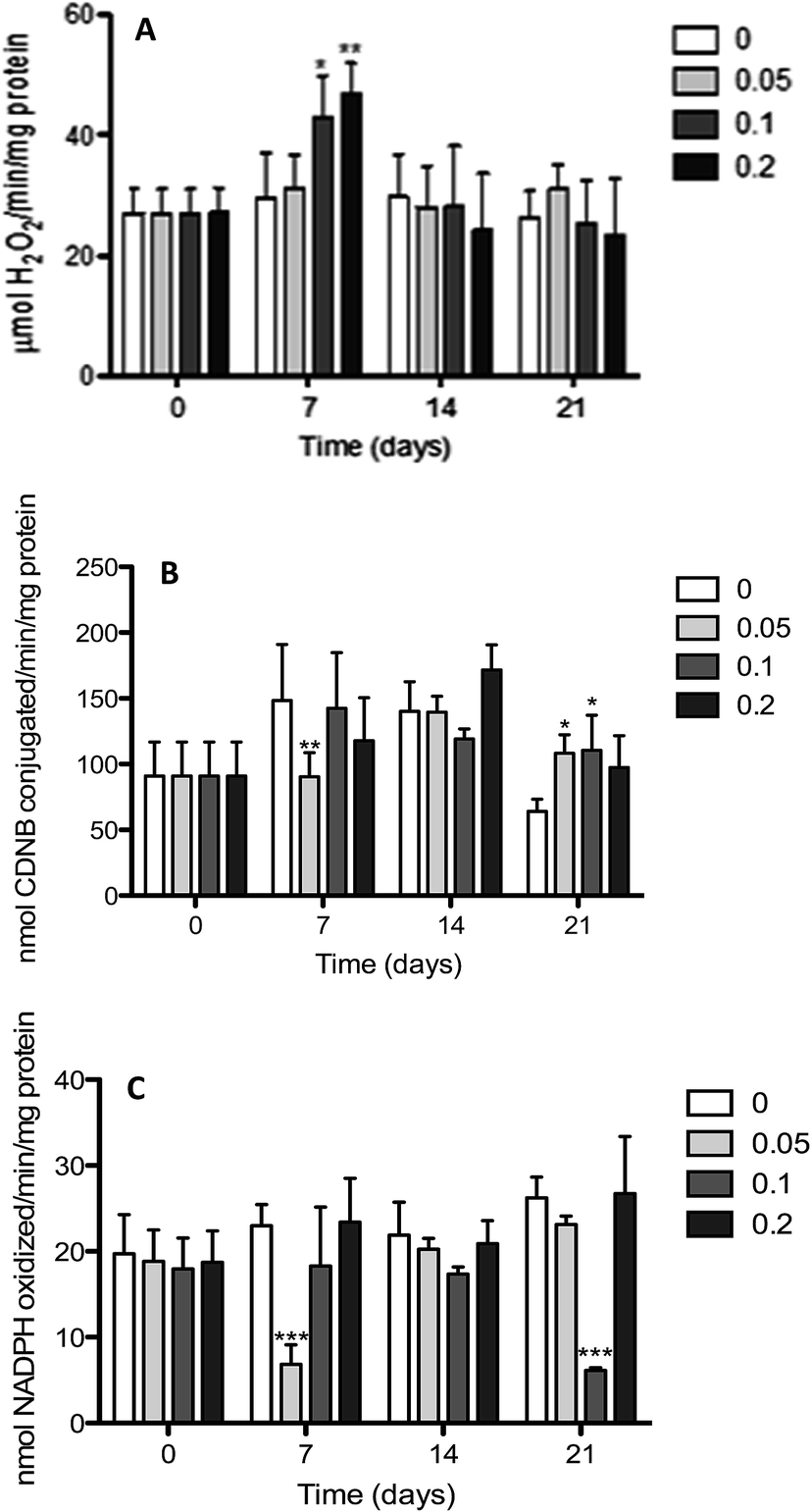 | ||
| Fig. 2 Gill antioxidant activities (means ± SD) in Nile tilapia O. niloticus exposed to different concentrations of Red 195 (0.05; 0.1 and 0.2 mg L−1). The activity levels of treated fish were compared with control group in each sampling day. Data are means ± SD of six independent experiments. *Values statistically different from control p < 0.05; **values statistically different from control p < 0.01; ***values statistically different from control p < 0.005. | ||
Furthermore, the results revealed that, compared to the control, the CAT activity in the gills (Fig. 2A) underwent a significant increase after 7 days of exposure to 0.1 and 0.2 mg L−1 of Red 195. No statistically significant differences in CAT activity were, however, observed for longer periods of exposure (14 and 21 days) with all concentrations (0.05, 0.1 and 0.2 mg L−1) when compared to the control. Further results showed an unexpectedly significant decrease in GST activity after 7 days of exposure to the lowest concentration of Red 195 (Fig. 2B). While GST activity displayed no statistically significant differences after 14 days, they underwent a significant increase after 21 days exposure to 0.05 and 0.1 mg L−1 of Red 195. Interestingly, the profile displayed by GR activity after 7 and 14 days of exposure was similar to the one observed for GST activity (Fig. 2C). After 21 days of exposure, however, the 0.1 mg L−1 concentration was noted induce a significant decrease in GR activity.
After 30 days of depuration, the antioxidant enzyme activities in the liver and gills did not show significant differences when compared to the ones observed after 21 days of exposure (data not shown).
3.2. Histology
The histological observations of control O. niloticus liver tissues (Fig. 3A and B) showed normal architecture, with the hepatocytes exhibiting a quite homogeneous cytoplasm and a large central or subcentral spherical nucleus. The liver of the fish exposed to Red 195 (Fig. 3C–F) showed several histopathological changes whose severity was noted to increase with the increase of Red 195 concentration and time of exposure. The hepatic parenchyma of fish exposed to 0.1 mg L−1 of Red 195 showed lower eosinophilia and some degree of cytoplasm vacuolization (Fig. 3C), which clearly increased with the time of exposure. The disruption of the endothelial lining was also evident after 7 days of exposure (Fig. 3C), becoming more prevalent after 21 days of exposure. An apparent presence of metabolic zonation was observed, with the vacuolization of hepatocyte being farthest from the afferent vascular area (Fig. 3D). Necrotic cells appeared in the periphery of vascular regions, and a rupture was observed in membrane of hepatocytes. These histopathological changes were more clearly evident after long periods of exposure (21 days), being accompanied by a higher prevalence of macrophage aggregates. However, the hepatic parenchyma of the fish exposed to 0.2 mg L−1 of Red 195 showed reduced eosinophilia and expansion of cytoplasmic vacuolization after both 7 and 21 days of exposure (Fig. 3E and F). The endothelial lining displayed a higher degree of rupture, with high levels of necrotic cells in the vascular periphery. A rupture in the hepatocytes membrane was also observed. Fig. 3G and H show the liver tissues from the recovery group (RG) of fish previously exposed to 0.1 and 0.2 mg L−1 of Red 195, respectively. Although some degree of hepatocyte cytoplasm vacuolization was observed, there were signs of regeneration. In general, an improvement in the hepatic parenchyma structure was detected for the RG. In some fish, the liver showed normal architecture, and the hepatocytes exhibited a quite homogeneous basophilic cytoplasm.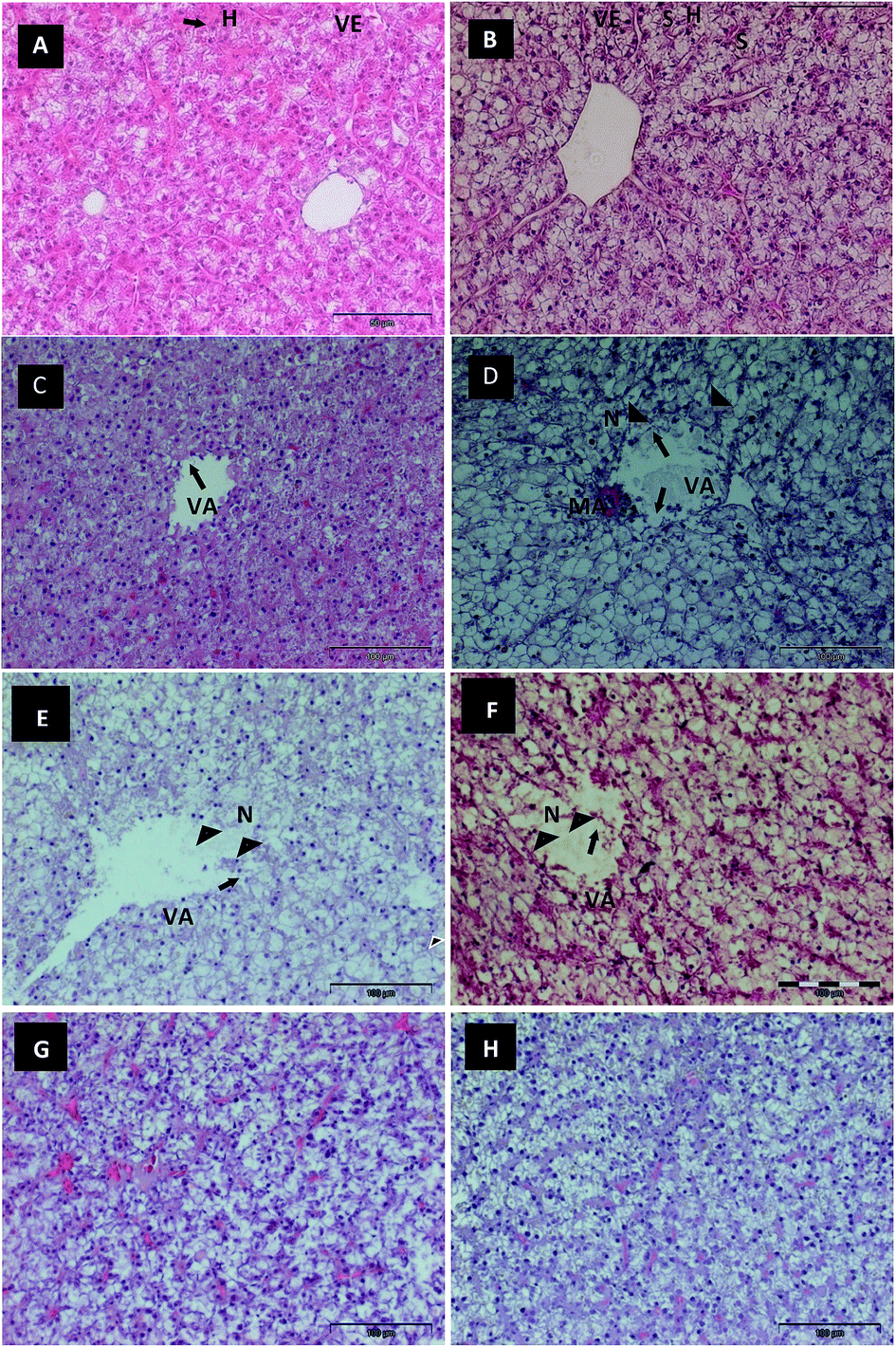 | ||
| Fig. 3 Histological images of Nile tilapia (O. niloticus) liver. (A and B) Control fish, showing a normal architecture, with the hepatocytes (H) exhibiting a quite homogeneous cytoplasm and a large central or subcentral spherical nucleus (arrow). S: sinusoid; VE: vein. (C–F) Liver from fish exposed to Red 195 dye; (C) (7th day, 0.1 mg L−1) showing cytoplasmic vacuolization (VA), disruption of the endothelial lining (arrow); (D) (21st day, 0.1 mg L−1) showing increased vacuolization (VA), severely disruption of the endothelial lining (arrow), a metabolic zonation, necrotic cells (N) and rupture of hepatocytes membrane (arrowhead), macrophage aggregates (MA); (E) (7th day, 0.2 mg L−1) and (F) (21st day, 0.2 mg L−1) showing higher eosinophilia reduction, increased cytoplasmic vacuolization, the endothelial lining showed a higher degree of rupture (arrow), with high level of necrotic cells and rupture of the hepatocytes membrane (arrowhead); (G and H) Recovering Group (RG) from fish liver previously exposed to 0.1 and 0.2 mg L−1 of Red 195 showing the signs of regeneration with the maintenance of hepatocyte cytoplasmic vacuolization and an improvement in the hepatic parenchyma structure. | ||
Further histological observations revealed that the gill tissues of the O. niloticus control group (Fig. 4A and B) showed normal architecture. The gill filaments consisted of a central axis with a connective tissue and venous sinus. The filaments were covered by a stratified epithelium, intersected by lamellae (Fig. 4A and B). The gills of the fish exposed to Red 195 (Fig. 4C–F) for 7 and 21 days showed histopathological changes, with the highest rates of severity being observed after long periods of exposure. The gills of fish exposed to 0.1 mg L−1 of Red 195 for 7 days (Fig. 4C) showed lamellar epithelium lifting associated with a high level of filament epithelium proliferation (FEP). After 21 days of exposure to the same Red 195 concentration (Fig. 4D), the filament epithelium proliferation induced, in some instances, lamellar fusion. Moreover, the results revealed the vasodilatation of the lamellar vascular axis and some necrotic cells in the deep region of the filament epithelium.
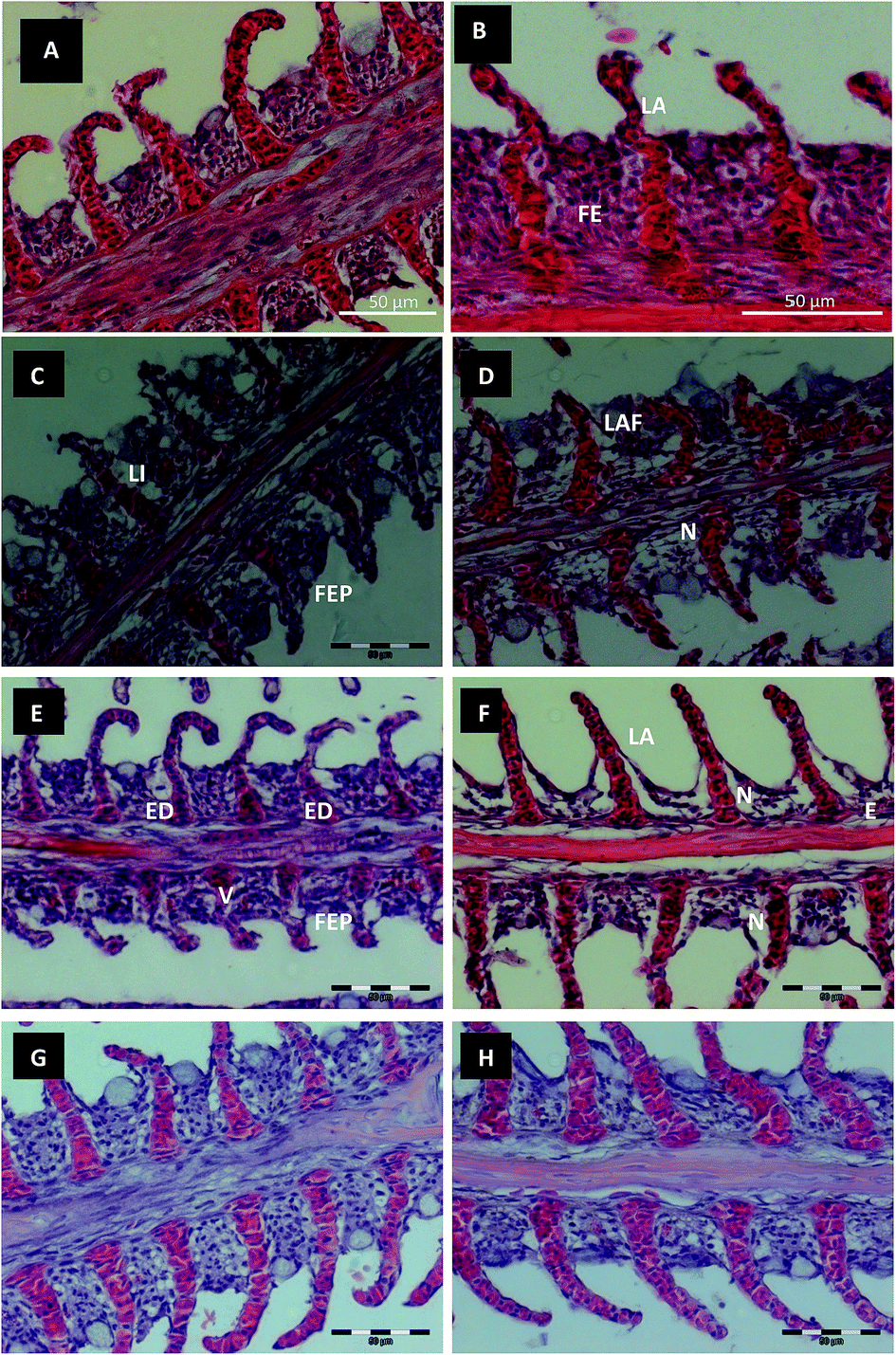 | ||
| Fig. 4 Histological images of Nile tilapia (O. niloticus) gills. (A and B) Control fish, showing a normal appearance of gill filament epithelium (FE) and lamellae (LA). (CT): connective tissue and (CVS): venous sinus. (C–F) Gills from exposed fish to Red 195 dye; (C) (7th day, 0.1 mg L−1) showing lifting (LI) of lamellar epithelium and filament epithelium proliferation (FEP); (D) (21st day, 0.1 mg L−1) present some necrotic cells (N) in the deep region of the filament epithelium, vasodilatation (V) and filament epithelium proliferation that in some instances conduced to lamellar fusion (LAF); (E) (7th day, 0.2 mg L−1) showing edema (ED), vascular disorders such as extreme vasodilatation (V) and proliferation of filament epithelium (FEP); (F) (21st day, 0.2 mg L−1) presence of necrotic cells (N) and lifting (LI) of filament and lamellar epithelium became evident, mainly in the deep region of the filament epithelium. (G and H) Recovering Group (RG) from fish gill previously exposed to 0.1 and 0.2 mg L−1 showing a clearly recover of the filament microarchitecture. In the majority of the fish, although the epithelial repair was not complete, clear signs of compensatory responses were evident. Bar = 50 μm. | ||
The histopathological changes observed for the gills of fish exposed to 0.2 mg L−1 of Red 195 were similar to those detected for the fish exposed to 0.1 mg L−1. The severity rate was, however, clearly higher after 21 days of exposure to the highest concentration of Red 195. Exposure to 0.2 mg L−1 for 7 days induced edema (Fig. 4E), which presumably induced filament lifting and lamellar epithelium (Fig. 4F). Vascular disorders, such as extreme vasodilatation and filament epithelium proliferation, were also observed. Furthermore, after 21 days of exposure, the presence of necrotic cells became more evident, mainly in the deep region of the filament epithelium (Fig. 4F).
After the depuration period, the gills of the fish previously exposed to 0.1 and 0.2 mg L−1 showed a clear recovery of the filament microarchitecture (Fig. 4G and H). In some fish, the gill epithelium recovered its normal structure. Although epithelial repair was not complete, clear signs of compensatory responses were still evident in most of the fish samples.
4. Discussion
Biochemical and histopathological responses have commonly been used as effective biomarkers for evaluating the effects of pollutants on the health and ecological status of aquatic biota. Accordingly, the present work aimed to evaluate the toxicity effects of Red 195 dye on O. niloticus gill and liver through the study of oxidative stress responses and histopathological changes. The results revealed that the oxidative stress-response enzymes evaluated in this study displayed different responses to this dye. The findings indicated that, compared to the control, the tilapia fish exposed to Red 195 dye for all the periods assayed in this work underwent a decrease of CAT activity (Fig. 1A) in the liver. This is in agreement with the results previously reported for other xenobiotics.35,40 The results showed, however, that CAT activity in the gills (Fig. 2A) increased after 7 days exposure, which is in agreement with the results previously observed for bivalves41–44 and fish exposed to several pollutants, including polycyclic aromatic hydrocarbons (PAHs) and polychlorinated biphenyls (PCBs).45,46Catalase activity is responsible for the conversion of hydrogen peroxide in water. However, the microsomal production of oxyradicales seems to be stimulated by a wide variety of xenobiotics including B(a)P, aroclor 1254,47 and HAPs,48 as well as pharmaceutical products, such as ubiprofen.49 The results of the present study revealed that longer periods of exposure to Red 195 dye induced a decrease in CAT activity. This could be attributed to the accumulation of hydrogen peroxide, which causes other damages to the tissue and cell. In fact, it is possible to have the production of other types of radicals via a Fenton reaction.
Furthermore, GST activity plays a central role both in the maintenance antioxidative defences50 and removal of pollutants by increasing their solubility in water through conjugation with GSH.51–53 The tilapia fish exposed to Red 195 displayed an increase in GST activity exclusively in the liver and only after 21 days of exposure to a high concentration 0.2 mg L−1. Conversely, the hepatic GST activity of tilapia species exposed to oxyfluorfen was previously reported to undergo an increase in the first week, but to exhibit a subsequent decrease with longer periods of exposure.24 However, an increase in hepatic GST activity was reported for fish exposed to PCBs, PAHs, and PCDDs (polychlorinated dibenzodioxins).54,55 Other studies showed that exposure to PCDD, PAHs, and organochlorine pesticides (OCPs) caused a decrease in enzymatic activity.56,57 In the gills, GST activity increased after 14 days of exposure to 0.2 mg L−1 of Red 195, although a significant increase was observed with all concentrations after 21 days of exposure. These observations suggested that the GST was able to eliminate the xenobiotic at this tissue. This result is in agreement with the experimental variations observed for the GST activity in the gills of Fulvia fragilis.44 However, a decrease in GST activity was previously reported for the gills of Donax trunculus originating from a Tunisian polluted site.58 Decreased GST activities have also been observed in other species of bivalves,59 gastropods60 and fish.61 Several xenobiotics can disrupt phase II enzymes of the cellular metabolic processes, inducing signal transduction events that can lead to various cellular and physiological responses. For pharmaceuticals, this takes place primarily through the various biotransformation reactions to be eliminated.49,62,63 Several studies have shown that the effects of GST may be caused by other pollutants, such as B(a)P and aroclor.47 In fact, different assumptions have been made about the metabolic pathways and mode of action of this dye on GST activity. The change observed in GST activity can reflect the ability of Red 195 to affect the enzymes involved in the maintenance of tissue redox balance.64 The changes of GST activity may correspond to the induction of a detoxifying enzyme in the gills of the fish exposed to the dye to respond to the invasion of the organism by the dye via its conjugation with a thiol group.65
The results showed an increase in hepatic GR activity for all periods of exposure and at all concentrations, except for the 0.2 mg L−1 concentration for 21 days of exposure. Peixoto et al. (2006)24 showed that GR activity increased in the liver of tilapia treated with oxyfluorfen at all sampling days. However, Matos et al., (2007)25 reported that carbamate treatment induced an increase in GR activity only after 14 days of treatment. While several studies showed that the exposure to different contaminants induced an increase in GR activity,66,67 other studies reported on a significant decrease in this activity, including the study on O. niloticus originating from a site contaminated with PCBs and HCHs (hexachlorocyclohexane).68 Unlike the case of GR activity in the liver, the results showed that GR activity in the gills decreased after 14 days treatment, with statistically significant values observed for the groups treated with Red 195 at concentrations of 0.1 and 0.2 mg L−1. This result suggested the activation of gill detoxification processes, presumably due to the presence of organic contaminants.69
The literature indicates that the adverse effects of pollutants on marine biota can be evaluated by histopathological alterations in fish organs, such as the liver and gills61 since they reflect the overall health of the fish population in the ecosystem.70 Morphological changes are generally indicative of irreversible damage. The results of the present study revealed that the most prominent histological alterations observed in the liver of Nile tilapia fish exposed to Red 195 were necrosis, hepatocytes cytoplasm vacuolization and macrophage aggregates. These histopathological changes were more clearly evident after long-term exposures, with a high level of necrotic cells. This result is in accordance with several reports in the literature.71,72 The higher rates of necrosis in fish exposed to different pollutants can be an internal response of the organism, as a consequence of the metabolic stress induced by the increase of xenobiotic availability during this period.72 This is consistent with the results observed for enzymatic activity. In fact, the results showed that CAT activity was inhibited, this reflecting the accumulation of H2O2 which, in turn, could lead to higher possibilities for the formation of other radicals and lipidic peroxidation. However, the fact that GR activity increased for all concentrations and with all periods of exposures suggested the presence of more GSSG. GST activity was, on the other hand, noted to undergo an increase or a decrease depending on the dose and duration of exposure. These variations highlighted the state of conjugation reactions of GSH. Exposure to Red 195 was noted to alter the GSH/GSSG ratio in tilapia. In fact, GST constitutes a complex family of proteins that play significant roles both in the normal cellular metabolism and the detoxification of a wide variety of xenobiotic compounds.
The vacuoles in the cytoplasm of the hepatocytes can contain lipids and glycogen, which are related to the normal metabolic function of the liver.73 The vacuolization of hepatocytes might indicate an imbalance between the rate of synthesis of substances in the parenchyma cells and the rate of their release into the circulation.74 Vacuole formation was considered as a cellular defense mechanism against substances injurious to hepatocytes, and this mechanism could be responsible for collecting the harmful elements and preventing them from interfering with the biological activities of these cells.75 An increase in the density of the macrophage aggregates is generally related to important hepatic lesions,76 such as degenerative and necrotic processes. Several studies have reported on other changes in the liver of O. niloticus and other fish species, resulting from exposure to different toxic chemicals.77–81 After the depuration period, the liver tissues of some fish samples previously exposed to Red 195 dye showed signs of regeneration. These results suggested that the toxic effects induced by Red 195 dye could be reversed after a depuration period of 30 days. In fact, the toxic effects of other toxicants, such as carbaryl, in O. niloticus species were previously shown to be relatively reversible within 15 days of exposure after stimulus withdrawal.25
Furthermore, the results revealed that the gill from the control O. niloticus fish displayed a typical structural organization. The histopathological changes of the gills from the fish exposed to Red 195 dye showed higher levels of severity after a long period of exposure, including lamellar epithelium lifting, epithelium proliferation, necrotic cells, lamellar axis vasodilation, fusion of lamellar, and edema in the filament. As an histological change, the lifting of lamellar epithelium could presumably be induced by the incidence of severe edema.82,83 Edema, together with the lifting of lamellar epithelium, could serve as a defense mechanism since epithelial separation from the lamellae increases the distance across which waterborne pollutants need to diffuse to reach the bloodstream.84
The cell proliferation observed with the thickening of gill filament epithelium in O. niloticus exposed to Red 195 dye could lead to the lamellar fusion. Similar results were previously reported in fish species exposed to other pollutants.85,86 Azevedo et al. (2013)72 showed that, in Cathorops spixii, if the pollutant pressure decreases, the lamellar fusions could be considered a reversible lesion with low severity. However, in cases of stress continuity, lamellar fusion could compromise the breathing of the fish and ultimately contribute to asphyxia or neoplasic events.72 Miranda et al. (2008)71 reported on similar results for fish species from aquatic systems under exposure to organochlorine compounds. The epithelial lifting, edema and lamellar fusion are defense mechanisms that reduce the branchial superficial area in contact with the external milieu. These mechanisms also increase the diffusion barrier to the pollutant.87
Lamellar axis vasodilatation was also reported for tilapia exposed to Red 195. Garcia-Santos et al. (2006)88 reported that this lesion could induce changes in the normal structure of pillar cells, with a consequent loss of their support function, which could presumably be responsible for the emergence of lamellar aneurysms in fish.
The cellular damages observed in the gills can adversely affect the gas exchange and ionic regulation processes, as a consequence of the increased distance between water and blood due to epithelial lifting.89
Similar histological alterations in the gills were previously observed in some fish species due to the exposure to different types of pollutants, including drugs.83 Accordingly, this could suggest that alterations are non-specific and can be induced by different types of contaminants.
The gill epithelium of the recovery group of fish previously exposed to Red 195 showed a relatively normal structure. Although the epithelial repair was not complete, clear signs of compensatory responses were evident. These results confirmed the findings observed for the liver of the O. niloticus fish.
Taken together, the results of the present study showed for the first time that low concentrations (0.05, 0.1, 0.2 mg L−1) of Red 195 dye could induce oxidative stress in O. niloticus. The oxidative stress was demonstrated by biochemical and histological changes in the liver and gill of tilapia. Biological responses were proved by increasing or inhibiting the activities of some antioxidant enzymes (CAT, GST and GR) and by histopathological changes observed in tilapia exposed to Red 195 dye. The results obtained for the recovery group suggested that the toxicity produced by Red 195 dye was, to some extent, reversible within 30 days after the removal of the stimulus. The biochemical and histological changes in the liver and gill were noted to depend on the concentration and period of exposure to Red 195. Overall, the results call for extreme caution to the adverse negative impacts of this dye on wild fish. Further studies are needed to explore the practical application of using bacterial species in the bioremediation of effluents containing this dye and some cases that involve the persistence of this dye in the water.
Conflict of interest
The authors declare that they have no competing interests.Acknowledgements
This study was co-funded by the Tunisian Ministry of Higher Education, Scientific Research and Information Technology and Communication and by the Center for the Research and Technology of Agro-Environmental and Biological Sciences, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal. Thanks are also to Dr Sharif Mohammad Shahidullah from Department of English, Faculty of Sciences and Arts in Balgarn, Bisha University, Saudi Arabia for his contribution in the correction of English language.References
- H. Zollinger, Color chemistry, syntheses, properties and applications of organic dyes and pigments, VCH, 1987 Search PubMed.
- P. Cooper, Colour in dyestuff effluent, the society of dyers and colourists, Aden, Oxford, 1995 Search PubMed.
- U. Pagga and D. Brown, The degradation of dyestuffs: Part II. Behaviour of dyestuffs in aerobic biodegradation tests, Chemosphere, 1986, 15(4), 479–491 CrossRef CAS.
- DEPA (Danish Environmental Protection Agency), Survey of azo-colorants in Denmark, toxicity and fate of azo dyes, 2000.
- C. Bauer, P. Jacques and A. Kalt, Photooxidation of an azo dye induced by visible light incident on the surface of TiO2, J. Photochem. Photobiol., A, 2001, 140, 87–92 CrossRef CAS.
- H. Ben Mansour, D. Corroler, D. Barillier, K. Ghedira, L. Chekir and R. Mosrati, Evaluation of genotoxicity and pro-oxidant effect of the azo dyes: acids yellow 17, violet 7 and orange 52, and of their degradation products by Pseudomonas putida mt-2, Food Chem. Toxicol., 2007, 45(9), 1670–1677 CrossRef CAS PubMed.
- H. Ben Mansour, D. Barillier, D. Corroler, K. Ghedira, L. Chekir-Ghedira and R. Mosrati, In vitro study of DNA damage induced by acid orange 52 and its biodegradation derivatives, Environ. Toxicol. Chem., 2009, 28(3), 489–495 CrossRef CAS PubMed.
- H. Ben Mansour, R. Mosrati, D. Corroler, K. Ghedira, D. Barillier and L. Chekir, In vitro mutagenicity of Acid Violet 7 and its degradation products by Pseudomonas putida mt-2: Correlation with chemical structures, Environ. Toxicol. Pharmacol., 2009, 27, 231–236 CrossRef CAS PubMed.
- H. Ben Mansour, Y. Ayed-Ajmi, R. Mosrati, D. Corroler, K. Ghedira, D. Barillier and L. Chekir-Ghedira, Acid violet 7 and its biodegradation products induce chromosome aberrations, lipid peroxidation, and cholinesterase inhibition in mouse bone marrow, Environ. Sci. Pollut. Res. Int., 2010, 17(7), 1371–1378 CrossRef CAS PubMed.
- K. Al-Sabti, Chlorotriazine Reactive Azo Red 120 Textile Dye Induces Micronuclei in Fish, Ecotoxicol. Environ. Saf., 2000, 47(2), 149–155 CrossRef CAS PubMed.
- M. Sumathi, K. Kalaiselvi, M. Palanivel and P. Rajaguru, Genotoxicity of Textile Dye Effluent on Fish (Cyprinus carpio) Measured Using the Comet Assay, Bull. Environ. Contam. Toxicol., 2001, 66, 407–414 CrossRef CAS.
- J. M. Law, Issues related to the use of fish models in toxicologic pathology: session introduction, Toxicol. Pathol., 2003, 31, 49–52 CAS.
- T. W. Moon, P. J. Walsh and T. P. Mommsen, Fish hepatocytes: a model metabolic system, Can. J. Fish. Aquat. Sci., 1985, 42, 1772–1782 CrossRef CAS.
- R. Triebskorn, H. R. Köhler, W. Honnen, M. Schramm and S. M. Adams, Induction of heat shock proteins, changes in liver ultrastructure, and alterations of fish behaviour: are these biomarkers related and are they useful to reflect the state of pollution in the field?, J. Aquat. Ecosyst. Stress Recovery, 1997, 6, 57–73 CrossRef CAS.
- S. Barhoumi, I. Messaoudi, T. Deli, K. Said and A. Kerkeni, Cadmium bioaccumulation in three benthic fish species, Salaria basilica, Zosterisessor ophiocephalus and Solea vulgaris collected from the Gulf of Gabes in Tunisia, J. Environ. Sci., 2009, 21, 980–984 CrossRef CAS.
- M. S. Shailaja and C. D. D'Silva, Evaluation of impact of PAH on a tropical fish, Oreochromis mossambicus using multiple biomarkers, Chemosphere, 2003, 53, 835–841 CrossRef CAS.
- D. E. Hinton and D. J. Laurén, Liver structural alterations accompanying chronic toxicity in fishes: potential biomarkers of exposure, in Biomarkers of Environmental Contamination, ed. J. F. McCarthy and L. R. Shugart, Lewis, Boca Raton, 1990, pp. 17–57 Search PubMed.
- S. Biagianti-Risbourg, Les perturbations (ultra) structurales du foie des poissons utilisées comme biomarqueurs de la qualité sanitaire des milieux aquatiques, in Utilisation de Biomarqueurs en Ecotoxicologie, Aspects Fondamentaux, ed. L. Lagadic, T. Caquet, J. C. Amiard and F. Ramade, Masson Pub., Paris, 1997, pp. 55–391 Search PubMed.
- R. T. Di Giulio, P. C. Washburn, R. J. Wenning, G. W. Winston and C. S. Jewell, Biochemical responses in aquatic animals: a review of oxidative stress, Environ. Toxicol. Chem., 1989, 8, 1103–1123 CrossRef CAS PubMed.
- D. Bagchi, M. Bagchi, E. A. Hassoun and S. J. Stohs, In vitro and in vivo generation of reactive oxygen species, DNA damage and lactate dehydrogenase leakage by selected pesticides, Toxicology, 1995, 104, 129–140 CrossRef CAS.
- A. Orbea, M. Ortiz-Zarragoitia, M. Solé, C. Porte and M. P. Cajaraville, Antioxidant enzymes and peroxisome proliferation in relation to contaminant body bordens of PAHs and PCBs in bivalve molluscs, crabs and fish from the Urdaibai and Plentzia estuaries (Bay of Biscay), Aquat. Toxicol., 2002, 58, 75–98 CrossRef CAS.
- F. Regoli, G. Frenzilli, R. Bochetti, F. Annarumma, V. Scarcelli, D. Fattorini and M. Nigro, Time-course variations of oxyradical metabolism, DNA integrity and lysosomal stability in mussels, Mytilus galloprovincialis, during a filed translocation experiment, Aquat. Toxicol., 2004, 68, 167–178 CrossRef CAS PubMed.
- A. A. Fontainhas-Fernandes, Tilapia production, in Aquaculture Handbook, ed. M. A. Reis-Henriques, 1998, pp. 135–150 Search PubMed.
- F. Peixoto, D. Alves-Fernandes, D. Santos and A. Fontaínhas-Fernandes, Toxicological effects of oxyfluorfen on oxidative stress enzymes in tilapia Oreochromis niloticus, Pestic. Biochem. Physiol., 2006, 85, 91–96 CrossRef CAS PubMed.
- P. Matos, A. Fontaınhas-Fernandes, F. Peixoto, J. Carrola and E. Rocha, Biochemical and histological hepatic changes of Nile tilapia Oreochromis niloticus exposed to carbaryl, Pestic. Biochem. Physiol., 2007, 89, 73–80 CrossRef CAS PubMed.
- S. A. Mansour and A. H. Mossa, Oxidative damage, biochemical and histopathological alteration in rat exposed to chlorpyrifos and the role of zinc as antioxidant, Pestic. Biochem. Physiol., 2010, 96, 14–23 CrossRef CAS PubMed.
- S. A. Mansour and A. H. Mossa, Lipid peroxidation and oxidative stress in rat erythrocytes induced by chlorpyrifos and the protective effect of zinc, Pestic. Biochem. Physiol., 2009, 93, 34–39 CrossRef CAS PubMed.
- F. Mohamed, L. Senarathna, A. Percy, M. Abeyewardene, G. Eaglesham, R. Cheng, S. Azher, A. Hittarage, W. Dissanayake, M. H. Sheriff, W. Davies, N. A. Buckley and M. Eddleston, Acute human self-poisoning with the N-phenylpyrazole insecticide fipronil – a GABAA-gated chloride channel blocker, J. Toxicol., Clin. Toxicol., 2004, 42, 955–963 CrossRef CAS PubMed.
- B. D. Banerjee, V. Seth, A. Bhattacharya, S. T. Pasha and A. K. Chakraborty, Biochemical effects of some pesticides on lipid peroxidation and free radical scavengers, Toxicol. Lett., 1999, 107, 33–47 CrossRef CAS.
- K. R. Tukhtaev, S. K. Tulemetov, N. B. Zokirova, N. K. Tukhtaev, M. R. Tillabaev, O. K. Amirullaev, O. O. Yarieva and A. N. Otajonova, Prolonged exposure of low doses of fipronil causes oxidative stress in pregnant rats and their offspring, Int. J. Toxicol., 2013, 10, 1–11 Search PubMed.
- N. Deb and S. Das, Chlorpyrifos Toxicity in Fish: a Review, Curr. World Environ., 2013, 8(1), 77–84 CAS.
- D. Ali, N. S. Nagpure, S. Kumar, R. Kumar, B. Kushwaha and W. S. Lakra, Assessment of genotoxic and mutagenic effects of chlorpyrifos in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline single-cell gel electrophoresis, Food Chem. Toxicol., 2009, 47, 650–656 CrossRef CAS PubMed.
- M. M. Abdel-Daim, N. K. Abdelkhalek and A. M. Hassan, Antagonistic activity of dietary allicin against deltamethrin-induced oxidative damage in freshwater Nile tilapia; Oreochromis niloticus, Ecotoxicol. Environ. Saf., 2015, 111, 146–152 CrossRef CAS PubMed.
- N. K. Abdelkhalek, E. W. Ghazy and M. M. Abdel-Daim, Pharmacodynamic interaction of Spirulina platensis and deltamethrin in freshwater fish Nile tilapia, Oreochromis niloticus: impact on lipid peroxidation and oxidative stress, Environ. Sci. Pollut. Res. Int., 2015, 22(4), 3023–3031 CrossRef CAS PubMed.
- A. A. Fontainhas-Fernandes, E. Gomes, M. A. Reis-Henriques and J. Coimbra, Replacement of fish meal by plant proteins in the diet of tilapia (Oreochromis niloticus): digestibility and growth performance, Aquacult. Int., 1999, 7, 57–67 CrossRef CAS.
- L. A. Del Rio, M. G. Ortega, A. L. Lopez and J. L. Gorge, A more sensitive modification of the catalase assay with the Clark oxygen electrode. Application to the kinetic study of the pea leaf enzyme, Anal. Biochem., 1977, 80, 409–415 CrossRef CAS.
- I. Carlberg and B. Mannervik, Glutathione reductase levels in rat brain, J. Biol. Chem., 1975, 250, 5375–5380 Search PubMed.
- P. J. Hatton, D. Dixon, D. J. Cole and R. Edwards, Glutathione transferase activities and herbicide selectivity in maize and associated weed species, Pestic. Sci., 1996, 46, 267–275 CrossRef CAS.
- M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding, Anal. Biochem., 1976, 72, 248–254 CrossRef CAS.
- H. Liu, W. Wang, J. Zhang and X. Wang, Effects of copper and its ethylenediaminetetraacetate complex on antioxidants defences of the gold-fish Carrasius auratus, Ecotoxicol. Environ. Saf., 2006, 650, 350–354 CrossRef PubMed.
- M. Dellali, M. Gnassia-Barelli, M. Romeo and P. Aissa, The use of acetylcholinesterase activity in Ruditapes decussatus and Mytilus galloprovincialis in the biomonitoring of Bizerta lagoon, Comp. Biochem. Physiol., Part C: Toxicol. Pharmacol., 2001, 130, 227–235 CrossRef CAS.
- M. Dellali, M. Romeo, M. Gnassia-Barelli and P. Aissa, A multivariate data analysis of the clam Ruditapes decussatus as sentinel organism of the Bizerta lagoon (Tunisia), Water, Air, Soil Pollut., 2004, 44, 131–156 CrossRef.
- A. Khessiba, M. Romeo and P. Aissa, Effects of some environmental parameters on catalase activity measured in the mussel (Mytilus galloprovincialis) exposed to lindane, Environ. Pollut., 2005, 133, 275–281 CrossRef CAS PubMed.
- N. Mahmoud, M. Dellali, M. El Bour, P. Aissa and E. Mahmoudi, The use of Fulvia fragilis (Mollusca: Cardiidae) in the biomonitoring of Bizerta lagoon: a multimarkers approach, Ecol. Indic., 2010, 10, 696–702 CrossRef CAS PubMed.
- D. R. Livingstone, P. Lemaire, A. Mathews, L. Peters, D. Bucke and R. J. Law, Pro-oxidant, antioxidant and 7-ethoxyresoruWn O-deethylase (EROD) activity responses in liver of dab (Limanda limanda) exposed to sediment contaminated with hydrocarbons and other chemicals, Mar. Pollut. Bull., 1993, 26, 602–606 CrossRef CAS.
- E. Stephensen, J. Svavarsson, J. Sturve, G. Ericon, M. Adolfson-Erici and L. Förlin, Biochemical indicators of pollution exposure in shorthorn sculpin (Myoxocephalus scorpius), caught in four harbours on the south-west coast of Iceland, Aquat. Toxicol., 2000, 48, 431–442 CrossRef CAS.
- C. C. C. Cheung, W. H. L. Siu, B. J. Richardson, S. B. D. Luca-Abbott and P. K. S. Lam, Antioxidant responses to benzo[a]pyrene and Aroclor 1254 exposure in the green-lipped mussel, Perna viridis, Environ. Pollut., 2004, 128, 393–403 CrossRef CAS PubMed.
- C. Porte, M. Sole, J. Albaiges and D. R. Livingstone, Responses of mixed-function oxygenases and antioxidant enzyme system of Mytilus sp. to organic pollution, Comp. Biochem. Physiol., Part C: Pharmacol., Toxicol. Endocrinol., 1991, 100, 183–186 CrossRef CAS.
- M. Gonzalez-Rey and M. J. Bebianno, Does non-steroidal anti-inflammatory (NSAID) Ibuprofen induces antioxidant stress and endocrine disruption in mussel Mytilus galloprovincialis, Environ. Toxicol. Pharmacol., 2012, 33(1), 361–371 CrossRef CAS PubMed.
- O. Hajime, K. Ozaki and H. Yoshikawa, Identification of cytochrome P450 and glutathione-S-transferase genes preferentially expressed in chemosensory organs of the swallowtail butterfly, Papilio xuthus L, Insect Biochem. Mol. Biol., 2005, 8, 837–846 Search PubMed.
- E. D. Goldberg and K. K. Bertine, Beyond the mussel watch-new directions for monitoring marine pollution, Sci. Total Environ., 2000, 247, 165–174 CrossRef CAS.
- G. L. Pennec and M. L. Pennec, Induction of glutathione-S-transferases in primary cultured digestive gland acini from the mollusc bivalve Pecten maximus (L.): application of a new cellular model in biomonitoring studies, Aquat. Toxicol., 2003, 64, 131–142 CrossRef.
- S. M. Moreiro and L. Guilhmino, The use of Mytilus galloprovincialis acethylesterase and glutathione S-transferase activities as biomarkers of environmental contamination along the northwest Portuguese coast, Environ. Monit. Assess., 2005, 105, 309–325 CrossRef.
- L. Förlin, S. Blom, M. Celander and J. Sturve, Effects on UDP glucuronosyl transferase, glutathione transferase, DT-diaphorase and glutathione reductase activities in rainbow trout liver after long-term exposure to PCB, Mar. Environ. Res., 1996, 42, 213–216 CrossRef.
- J. D. Chen, F. H. Yew and G. C. Li, Thermal adaptation and heat shock response of tilapia ovary cells, J. Cell. Physiol., 1998, 134, 189–199 CrossRef PubMed.
- J. R. Pedrajas, J. Peinado and J. Lopez-Barea, Oxidative stress in fish exposed to model xenobiotics, oxidatively modified forms of Cu, Zn superoxide dismutase as potential biomarkers, Chem.-Biol. Interact., 1995, 98, 267–282 CrossRef CAS.
- P. Lemaire and L. Förlin, Livingstone D. Responses of hepatic biotransformation and antioxidant enzymes to CYP1A-inducers (3-methylcholantrene, B-naphthoflavone) in sea bass (Dicentrarchus labrax), dab (Limanda limanda) and rainbow trout (Oncorhynchus mykiss), Aquat. Toxicol., 1996, 36, 141–160 CrossRef CAS.
- S. Tlili, I. Métais, H. Boussetta and C. Mouneyrac, Linking changes at sub-individual and population levels in Donax trunculus: Assessment of marine stress, Chemosphere, 2010, 8, 692–700 CrossRef PubMed.
- B. J. Richardson, E. Mak, S. B. De Luca-Abbott, M. Martin, K. McClellan and P. K. S. Lam, Antioxidant responses to polycyclic aromatic hydrocarbons and organochlorine pesticides in green lipped mussels (Perna viridis): do mussels ‘‘integrate” biomarker responses?, Mar. Pollut. Bull., 2008, 57, 503–514 CrossRef CAS PubMed.
- M. Roméo, S. Gharbi-Bouraoui, M. Gnassia-Barelli, M. Dellali and P. Aissa, Responses of Hexaplex (Murex) trunculus to selected pollutants, Sci. Total Environ., 2006, 359, 135–144 CrossRef PubMed.
- H. van der Oost, J. Beyer and P. E. Vermeulen, Fish bioaccumulation and biomarkers in environmental risk assessment: a review, Environ. Toxicol. Pharmacol., 2003, 13, 57–149 CrossRef.
- M. Gonzalez-Rey and M. J. Bebianno, Does selective serotonin reuptake inhibitor (SSRI) fluoxetine affect mussel Mytilus galloprovincialis, Environ. Pollut., 2013, 174, 200–209 CrossRef PubMed.
- M. Gonzalez-Rey, J. J. Mattos, C. E. Piazza, A. C. D. Bainy and M. J. Bebianno, Effects of active pharmaceutical ingredients mixtures in mussel Mytilus galloprovincialis, Aquat. Toxicol., 2014, 153, 12–26 CrossRef CAS PubMed.
- L. Canesia, R. Fabbri, G. Gallo, D. Vallotto, A. Marcomini and G. Pojana, Biomarkers in Mytilus galloprovincialis exposed to suspensions of selected nanoparticles (Nano carbon black, C60 fullerene, Nano-TiO2, Nano-SiO2), Aquat. Toxicol., 2010, 100, 168–177 CrossRef PubMed.
- S. Kaur and A. Kaur, Variability in antioxidant/detoxification enzymes of Labeo rohita exposed to an azo dye, acid black (AB), Comp. Biochem. Physiol., Part C: Toxicol. Pharmacol., 2015, 167, 108–116 CrossRef CAS PubMed.
- S. Rajeshkumar, J. Mini and N. Munuswamy, Effects of heavy metals on antioxidants and expression of HSP70 in different tissues of Milk fish (Chanos chanos) of Kaattuppalli Island, Chennai, India, Ecotoxicol. Environ. Saf., 2013, 98, 8–18 CrossRef CAS PubMed.
- S. Yang, F. Xu, B. Zheng, F. Wu and S. Wang, Multibiomarker responses upon exposure to tetrabromobisphenol A in the freshwater fish Carassius auratus, Aquat. Toxicol., 2013, 142–143, 248–256 CrossRef CAS PubMed.
- A. C. D. Bayne, E. Saito, P. S. M. Carvalho and V. B. C. Junqueira, Oxidative stress in gill, erythrocytes, liver and kidney of Nile tilapia (Oreochromis niloticus) from a polluted site, Aquat. Toxicol., 1996, 34, 151–162 CrossRef.
- I. Ahmad, M. Pacheco and A. M. Santos, Anguilla anguilla L. oxidative stress biomarkers: an in situ study of freshwater wetland ecosystem (Pateira de Fermentelos, Portugal), Chemosphere, 2006, 65, 952–962 CrossRef CAS PubMed.
- Z. Khoshnood, S. Khodabandeh, S. Mosafer and R. Khoshnood, Effects of Cortisol on Gill Chloride Cells in Persian Sturgeon, Acipenser persicus, Fry, Yakhteh, 2010, 11(4), 424–431 Search PubMed.
- A. L. Miranda, H. Roche, M. A. F. Randi, M. L. Menezes and C. A. Oliveira Ribeiro, Bioaccumulation of chlorinated pesticides and PCBs in the tropical freshwater fish Hoplias malabaricus: histopathological, physiological and immunological findings, Environ. Int., 2008, 34(7), 939–949 CrossRef CAS PubMed.
- J. S. Azevedo, E. S. Braga, H. C. Silva de Assis and C. A. Oliveira Ribeiro, Biochemical changes in the liver and gill of Cathorops spixii collected seasonally in two Brazilian estuaries under varying influences of anthropogenic activities, Ecotoxicol. Environ. Saf., 2013, 96, 220–230 CrossRef CAS PubMed.
- M. M. P. Camargo and C. B. R. Martinez, Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream, Neotropical Ichthyology, 2007, 5(3), 327–336 CrossRef.
- W. H. Gingerich, Hepatic Toxicology of Fishes, in Aquatic Toxicology, ed. L. J. Weber, Raven Press, New York, 1982, pp. 55–105 Search PubMed.
- A. Mollendorf, Cytology and cell physiology, Academic press, New York, 3rd edn, 1973 Search PubMed.
- M. Pacheco and M. A. Santos, Biotransformation, genotoxic and histopathological effects of environmental contaminants in European eel, Anguilla anguilla L, Ecotoxicol. Environ. Saf., 2002, 53, 331–347 CrossRef CAS.
- E. Olojo, K. Olurin, G. Mbaka and A. Oluwemimo, Histopathology of the Gill and Liver Tissues of the African Catfish Clarias gariepinus Exposed to Lead, Afr. J. Biotechnol., 2005, 4(1), 117–122 CAS.
- A. Figueiredo-Fernandes, A. Fontaínhas-Fernandes, R. A. F. Monteiro, M. A. Reis-Henriques and E. Rocha, Effects of the fungicide mancozeb in the liver structure of Nile tilapia, Oreochromis niloticus-assessment and quantification of induced cytological changes using qualitative histopathology and the stereological point-sampled intercept method, Bull. Environ. Contam. Toxicol., 2006, 76(2), 249–255 CrossRef CAS PubMed.
- A. Figueiredo-Fernandes, A. Fontaínhas-Fernandes, F. Peixoto, E. Rocha and M. A. Reis-Henriques, Effect of paraquat on oxidative stress enzymes in tilapia Oreochromis niloticus at two levels of temperature, Pestic. Biochem. Physiol., 2006, 85, 97–103 CrossRef CAS PubMed.
- A. G. M. Osman, A. M. Abd El Reheem, K. Y. Abuel-Fadl and A. G. GadEl-Rab, Enzymatic and Histopathologic Biomarkers as Indicators of Aquatic Pollution in Fishes, Nat. Sci., 2010, 2(11), 1302–1311 CAS.
- A. A. Hadi and S. F. Alwan, Histopathological changes in gills, liver and kidney of fresh water fish, Tilapia zillii, exposed to aluminum, Int. J. Pharm. Life Sci., 2012, 3(11), 2071–2081 CAS.
- E. F. Pane, A. Haque and C. M. Wood, Mechanistic analysis of acute, Ni induced respiratory toxicity in the rainbow trout (Oncorhynchus mykiss): an exclusively branchial phenomenon, Aquat. Toxicol., 2004, 69, 11–24 CrossRef CAS PubMed.
- J. Schwaiger, H. Ferling, U. Mallow, H. Wintermayr and R. D. Negele, Toxic effects of the non-steroidal anti-inflammatory drug diclofenac. Part I. Histopathological alterations and bioaccumulation in rainbow trout, Aquat. Toxicol., 2004, 68, 141–150 CrossRef CAS PubMed.
- J. M. Arellano, V. Storch and C. Sarasquete, Histological changes and copper accumulation in liver and gills of the Senegales sole, Solea senegalensis, Ecotoxicol. Environ. Saf., 1999, 44, 62–72 CrossRef CAS PubMed.
- M. R. van den Heuvel, M. Power, J. Richards, M. MacKinnon and D. G. Dixon, Disease and gill lesions in yellow perch (Perca flavescens) exposed to oil sands mining-associated water, Ecotoxicol. Environ. Saf., 2000, 46, 334–341 CrossRef CAS PubMed.
- M. Rosety-Rodríguez, F. J. Ordoñez, J. M. Rosety, I. Rosety, A. Ribelles and C. Carrasco, Morpho-histochemical changes in the gills of turbot, Scophthalmus maximus L., induced by sodium dodecyl sulfate, Ecotoxicol. Environ. Saf., 2002, 51, 223–228 CrossRef PubMed.
- D. van Heerden, A. Vosloo and M. Nikinmaa, Effects of short-term copper exposure on gill structure, methallothionein and hypoxia-inducible factor-1α (HIF-1α) levels in rainbow trout (Oncorhynchus mykiss), Aquat. Toxicol., 2004, 69, 271–280 CrossRef CAS PubMed.
- S. Garcia-Santos, A. Fontaínhas-Fernandes and J. M. Wilson, Cadmium tolerance in the Nile tilapia (Oreochromis niloticus) following acute exposure: assessment of some ionoregulatory parameters, Environ. Toxicol., 2006, 21(6), 33–46 CrossRef CAS PubMed.
- A. G. M. Osman, Biomarkers in Nile Tilapia Oreochromis niloticus niloticus (Linnaeus, 1758) to Assess the Impacts of River Nile Pollution: Bioaccumulation, Biochemical and Tissues Biomarkers, J. Environ. Prot., 2012, 3, 966–977 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2015 |