
Kinetic study of DNA hybridization on DNA-modified gold nanoparticles with engineered nano-interfaces†
Akari Takashima and
Motoi Oishi*
Division of Materials Science, Faculty of Pure and Applied Sciences, University of Tsukuba, 1-1-1 Tennoudai, Tsukuba, Ibaraki 305-8573, Japan. E-mail: oishi@ims.tsukuba.ac.jp; Tel: +81-29-853-5935
First published on 2nd September 2015
Abstract
Colorimetric DNA sensing systems using DNA-modified gold nanoparticles (GNPs) rely on DNA hybridization events on the GNP. We report a systematic study of DNA hybridization kinetics on GNPs with different surface modifications using probe DNA and different molecular weight PEGs. The DNA hybridization kinetics are extremely sensitive not only to probe DNA density but also to the molecular weight of PEG.
DNA-modified gold nanoparticles (GNPs) have attracted much attention in the field of colorimetric DNA sensing systems, since Mirkin's group first reported that a significant red-to-purple color change is easily visible to the naked eye when DNA-modified GNPs are cross-linked to form aggregates through hybridization of target DNA and probe DNA immobilized on the GNPs.1 On the basis of this distinctive feature, various colorimetric biosensing systems using GNPs have been developed.2,3 Recently, we have reported a new GNP-based colorimetric DNA sensing system using a combination of an enzyme-free click chemical ligation chain reaction on GNPs and a magnetic separation process.4 This sensing system relies on click chemical ligation5–7 between a dibenzocyclooctyne-containing probe DNA and an azide-containing probe DNA on a GNP through hybridization with target DNA to amplify both the target DNA and GNPs with ligated DNA by using a thermal cycler. After magnetic separation of the GNPs with ligated DNA, ultrasensitive detection (50 zM: several copies) of target DNA (RNA), with sensitivity comparable to that of polymerase chain reaction (PCR)-based techniques, could be attained.
DNA hybridization events between target DNA and probe DNA on a GNP are key steps for GNP-based sensing systems, but, surprisingly, little information is available about DNA hybridization on GNPs to date. For instance, although several reports have been published on the thermodynamic study of DNA hybridization on GNPs,8–10 these papers only provide information about the stability of the DNA duplex on GNPs at equilibrium. Conversely, kinetic study provides information about the hybridization rate, in addition to the hybridization efficiency (HE) before the hybridization equilibrium is reached. Compared to thermodynamic studies, kinetic studies will provide more significant insights into both signal generation and detection times of GNP-based sensing systems, because biosensors are required to detect target molecules within a finite period. To our knowledge, however, there has been only one reported kinetic study of DNA hybridization on GNPs. Zhao et al. pointed out that the rate constants for DNA hybridization on GNPs were about an order of magnitude smaller than those for the hybridization of free DNA in solution.11 Furthermore, kinetic studies of DNA hybridization between DNA-modified GNPs (aggregation) have been reported, but their rate constants and equilibrium constants are not well understood.12,13 Accordingly, a systematic study on the influence of nano-interface structures of GNPs on DNA hybridization kinetics remains desirable to improve the sensitivity and detection time of GNP-based sensing systems.
Herein, we report a systematic study of DNA hybridization kinetics on GNPs with engineered nano-interfaces (Fig. 1). To make quantitative comparisons, five types of GNPs with different surface modifications, two types of DNA-modified GNPs, and three types of DNA/poly(ethylene glycol) (PEG)-co-modified GNPs with the same probe DNA density and PEGs of different molecular weights, were prepared. The kinetic study of hybridization of target DNA on these GNPs was carried out using a fluorescence-based method for comparing DNA hybridization behavior on each GNP surface. This method enabled the determination of the HE and apparent hybridization rate constant (appkh). The DNA hybridization kinetics on GNPs are extremely sensitive, not only to surface charges but also to probe DNA density, as well as to the molecular weight of PEG. We believe that this study provides a method for predicting the hybridization rate and HE on GNPs, which is important information for the rational design of DNA-modified GNP-based sensing systems.
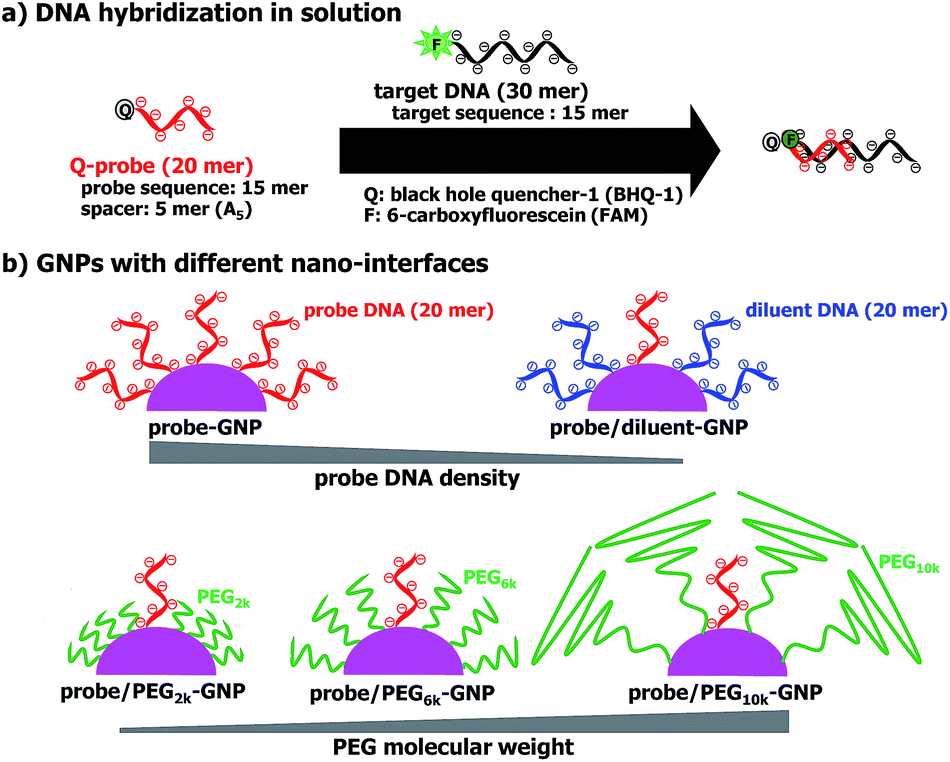 | ||
| Fig. 1 Schematic illustration of (a) DNA hybridization of target DNA and Q-probe in solution and (b) GNPs with different nano-interfaces. | ||
The probe DNA, diluent DNA, and target DNA sequences used in this study are listed in Table S1 (ESI†). To prevent lying orientation of the DNA strands on GNPs through Au–adenine interaction, we used dithiolated-probe DNA and dithiolated-diluent DNA to form preferentially standing orientation of DNA strands on GNPs through the two Au–S linkages. To clarify the influence of the probe DNA density on the hybridization of target DNA on GNPs, two types of DNA-modified GNPs were prepared. One had probe DNA composed of a spacer sequence (A5) and a half-complementary sequence (15 mer) to the target DNA (30 mer) (probe-GNP), and the other had both probe DNA and diluent DNA (A20) in a 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :8 ratio (probe/diluent-GNP). Additionally, three types of DNA/PEG-co-modified GNPs that had the same probe DNA density and PEGs with different molecular weights were prepared (probe/PEGx-GNPs, where subscript x denotes molecular weights of 2k, 6k, or 10k) to clarify the influence of particle charge and PEG molecular weight on the hybridization of DNA on GNPs. The average particle size and average number of DNA strands per particle of the prepared GNPs are listed in Table 1. Dynamic light scattering (DLS) measurements were carried out to determine the approximate length of the probe DNA on the GNPs. The lengths of the probe DNA and PEGs were estimated as half of the size increase after modifications. The probe DNA on GNPs was estimated to be approximately 7.0 nm in length, similar to values reported in the literature.14 DLS measurements of PEGx-GNPs without the probe DNA were also carried out to determine the approximate length of PEGs immobilized on the GNPs. (Table S3, ESI†). For PEG6k and PEG10k, the sizes of probe/PEGx-GNPs (x = 6k and 10k) were close to those of the respective PEGx-GNPs (x = 6k and 10k). However, the size of the probe/PEG2k-GNP (29.7 nm) was found to be intermediate to that of the probe-GNP (33.6 nm) and the PEG2k-GNP (25.3 nm), presumably due to the contribution of protruding probe DNA from the PEG2k layer.10 Consequently, PEG2k, PEG6k, and PEG10k on GNPs were estimated to be approximately 3.0, 4.5, and 9.0 nm in length, respectively. These values also show fair agreement with the literature.15 Therefore, the order of strand length at nano-interfaces is PEG10k > probe DNA (diluent DNA) > PEG6k > PEG2k, suggesting that the probe DNA protrudes from both the PEG6k and PEG2k layers while it is buried completely under the PEG10k layer.
:8 ratio (probe/diluent-GNP). Additionally, three types of DNA/PEG-co-modified GNPs that had the same probe DNA density and PEGs with different molecular weights were prepared (probe/PEGx-GNPs, where subscript x denotes molecular weights of 2k, 6k, or 10k) to clarify the influence of particle charge and PEG molecular weight on the hybridization of DNA on GNPs. The average particle size and average number of DNA strands per particle of the prepared GNPs are listed in Table 1. Dynamic light scattering (DLS) measurements were carried out to determine the approximate length of the probe DNA on the GNPs. The lengths of the probe DNA and PEGs were estimated as half of the size increase after modifications. The probe DNA on GNPs was estimated to be approximately 7.0 nm in length, similar to values reported in the literature.14 DLS measurements of PEGx-GNPs without the probe DNA were also carried out to determine the approximate length of PEGs immobilized on the GNPs. (Table S3, ESI†). For PEG6k and PEG10k, the sizes of probe/PEGx-GNPs (x = 6k and 10k) were close to those of the respective PEGx-GNPs (x = 6k and 10k). However, the size of the probe/PEG2k-GNP (29.7 nm) was found to be intermediate to that of the probe-GNP (33.6 nm) and the PEG2k-GNP (25.3 nm), presumably due to the contribution of protruding probe DNA from the PEG2k layer.10 Consequently, PEG2k, PEG6k, and PEG10k on GNPs were estimated to be approximately 3.0, 4.5, and 9.0 nm in length, respectively. These values also show fair agreement with the literature.15 Therefore, the order of strand length at nano-interfaces is PEG10k > probe DNA (diluent DNA) > PEG6k > PEG2k, suggesting that the probe DNA protrudes from both the PEG6k and PEG2k layers while it is buried completely under the PEG10k layer.
The average number of probe DNA strands per particle was estimated by using methods reported in the literature.4,16 The number of probe DNA strands for probe-GNP was 152 strands per particle (high density), whereas for other modified GNPs, the number could be adjusted to be 15–17 strands per particle (low density). Importantly, the difference in nano-interfaces between probe-GNP and probe/diluent-GNP was only the probe DNA density, while the total number of DNA strands (sum of probe and diluent) per particle was the same, indicating that the particle charge is also the same. Moreover, three probe/PEGx-GNPs will have lower particle charge than probe/diluent-GNP, since PEGs are nonionic while the diluent DNA is anionic, and the probe DNA density of probe/PEGx-GNPs is the same as that of probe/diluent-GNP.
DNA hybridization at the solid–liquid interface is affected by many factors such as DNA strand density (particle charge), DNA length, and polymer brush effect.17,18 Since these factors make the DNA hybridization kinetics at solid–liquid interfaces much more complicated than those for free DNA in solution, there are no reasonable fits to DNA hybridization kinetics on particles with the simple second-order kinetic model that had been adequate for the analysis of DNA hybridization in solution. Meanwhile, there are some reports on the determination of appkh at solid–liquid interfaces, including micro-particles, and planar substrates.19–24 These reports showed that fits to the Langmuir kinetic model that had been adequate for the analysis of adsorption of molecules on surfaces. To discuss DNA hybridization kinetics on GNPs using the Langmuir kinetic model, we maintained a low probe DNA density on probe/diluent-GNP and probe/PEGx-GNPs to reduce the effect of hybridized target DNA strands on further hybridization events with remaining probe DNA strands. Förster resonance energy transfer (FRET)-based method was chosen to monitor DNA hybridization in real time, because this method is commonly used for the observation of DNA hybridization, not only in solution but also at solid–liquid interfaces. In this study, target DNA labeled with a 6-carboxy-fluorescein (FAM) at the 3′-end and GNPs were used as a fluorescence dye and a quencher, respectively, because GNPs are well known for their ultrahigh fluorescence quenching ability. Additionally, probe DNA labeled with a black hole quencher-1 (BHQ-1) at the 5′-end (Q-probe) was used in control experiments to monitor DNA hybridization of free DNA in solution.
All DNA hybridizations were carried out using target DNA and probe DNA of the same concentration (7 nM) at 20 °C for 5400 s in 10 mM phosphate buffer (pH 7.0) containing 0.15 M NaCl and 0.1 wt% bovine serum albumin (BSA). Fig. 2 shows the time-dependent HE of the Q-probe and GNPs. The fluorescence intensity of the hybridization mixture containing hybridized target DNA on GNP and free target DNA at equilibrium was the same as that of the supernatant containing only free DNA after centrifugation of the hybridization mixtures. This indicates that FRET between GNPs and hybridized target DNA occurred quantitatively because the FAM dye of the hybridized target DNA is located on GNP surface within approximately 1.5 nm distance (A5 spacer). The time-dependent HE (%) of GNPs, therefore, which could be calculated by 100(1 − Ft/F0) as an eqn S(1) (ESI†), where Ft and F0 are the fluorescence intensity at t (s) and fluorescence intensity at initial time t = 0 (s), respectively. Although the HE of the Q-probe could not be determined simply by the change in fluorescence intensity, DNA hybridization of the Q-probe is assumed to occur quantitatively because the equilibrium (binding) constants (Keq) for DNA hybridization in solution are over 1010 (M−1) at low temperatures (T ≪ Tm). The HE at 5400 s, appkh, and apparent equilibrium (binding) constants (appKeq) are also listed in Table 2. The appkh and appKeq values for DNA hybridization on GNPs and in solution (Q-probe) were estimated from the Langmuir kinetic model-based fitting lines (Fig. 2 and see: ESI†) and the second-order kinetic model (Fig. S1, ESI†), respectively. As expected, all DNA hybridizations on GNPs were less efficient than that of Q-probe (free DNA) in solution. In particular, probe-GNP showed only 14% HE and a 25-fold decrease in appkh compared with the corresponding values of Q-probe in solution, and this fact is in accordance with the previous literature.11 Note that probe/diluent-GNP, with low probe DNA density, showed a slightly higher HE and appkh value than probe-GNP with high probe DNA density, even though the total number of DNA strands per particle (particle charge) is the same. Since the concentrations of target DNA and probe DNA in solution are the same (7 nM), particle concentrations of probe-GNP (2.8 × 1010 particles per mL) and probe/diluent-GNP (2.5 × 1011 particles per mL) are different. Based on the HE at 5400 s, the average number of hybridized target DNA strands per particle for probe-GNP (21 strands per particle) was greater than that for probe/diluent-GNP (3 strands per particle), and therefore the average distance between hybridized target DNA strands on probe-GNP (6.5 nm) was shorter than that on probe/diluent-GNP (17.3 nm). Mirkin et al. reported that an increase in the number of hybridized target DNA strands on GNPs makes further hybridization more difficult, due to increase in electrostatic repulsion.8 Indeed, the appKeq value of probe-GNP is slightly lower than that of probe/diluent-GNP (Table 2). These facts indicate that further hybridization of the target DNA to the remaining probe DNA strands on probe-GNP was inhibited by an overwhelming electrostatic repulsion and steric hindrance caused by the hybridized target DNA strands (Fig. 3a). In contrast to probe/diluent-GNP with low probe DNA density (2.4 × 1012 strands per cm2), the Langmuir kinetic model seems to be not suitable for DNA hybridization kinetic study on probe-GNP with high probe DNA density (2.2 × 1013 strands per cm2). Furthermore, the lower particle concentration of probe-GNP will result in a decrease in the collision frequency between probe-GNP and target DNA, resulting in inefficient DNA hybridization. Thus, the probe DNA density on GNPs was also found to be an important factor for the DNA hybridization kinetics on GNPs.
 | ||
| Fig. 2 Time-dependent hybridization efficiency (HE) of Q-probe and GNPs. The symbols and lines are the experimental data and the fitting based on the Langmuir kinetic model, respectively. Mean values and standard deviations were obtained from three or four independent experiments. | ||
| HE at 5400 s (%) | appkha (M−1 s−1) | appKeqa (M−1) | |
|---|---|---|---|
| a The appkh and appKeq values for DNA hybridization on GNPs were estimated by the Langmuir kinetic model.b The appkh and appKeq values for DNA hybridization of Q-probe in solution were estimated by the second-order kinetic model. | |||
| Q-probe (solution) | 98 ± 4 | (4.8 ± 0.8) × 105 b | (1.2 ± 2.0) × 1013 b |
| Probe-GNP | 14 ± 4 | (2.0 ± 0.7) × 104 | (2.3 ± 0.7) × 107 |
| Probe/diluent-GNP | 17 ± 2 | (3.6 ± 0.7) × 104 | (2.9 ± 0.4) × 107 |
| Probe/PEG2k-GNP | 45 ± 3 | (2.7 ± 0.4) × 105 | (1.1 ± 0.3) × 108 |
| Probe/PEG6k-GNP | 44 ± 4 | (2.0 ± 0.2) × 105 | (1.1 ± 0.1) × 108 |
| Probe/PEG10k-GNP | 37 ± 4 | (6.6 ± 0.7) × 104 | (1.0 ± 0.3) × 108 |
 | ||
| Fig. 3 Schematic illustration of (a) the change in the nano-interface structures of probe-GNP caused by hybridized target DNA and (b) the nucleation step for probe/PEGx-GNPs. The size of the arrows indicates the likelihood of the access of the target DNA to the probe DNA on GNPs. | ||
DNA hybridization on probe/PEGx-GNPs with both low probe DNA density and nonionic PEGs instead of anionic diluent DNA was also investigated. Note that probe/PEG10k-GNP showed a significantly higher HE and appkh value compared with those of probe/diluent-GNP, even though the probe DNA on probe/PEG10k-GNP is buried completely under the PEG10k layer. This indicates that electrostatic repulsion between target DNA and immobilized DNA on GNPs (particle charge) is a factor for DNA hybridization on GNPs. Importantly, further increase in appkh values and HE was observed for both probe/PEG2k-GNP and probe/PEG6k-GNP due to diminished steric hindrance of PEG chains on GNPs. Thus, it is remarkable that these appkh values of probe/PEG2k-GNP (2.7 × 105 M−1 s−1) and probe/PEG6k-GNP (2.0 × 105 M−1 s−1) are of the same order of magnitude as the appkh value of Q-probe (free DNA) in solution (4.8 × 105 M−1 s−1).
Based on these observations, we focus on the mechanism of DNA hybridization on GNPs. The DNA hybridization processes of free DNA in solution have been well studied and they involve two-step processes.25 The first step is the formation of a few base pairs between DNA strands to form a transient intermediate called a nucleus. The second step is the rapid formation of a complete duplex through the zipping of the remaining bases. Unlike fast zipping step of Q-probe (free DNA) in solution, the zipping step on GNP is likely to be slow due to electrostatic repulsion between the target DNA and the diluent DNA and steric hindrance of PEGs and the diluent DNA, leading to lower HE of probe/diluent-GNP and probe/PEGx-GNPs compared with that of Q-probe. The highest appkh of probe/PEG2k-GNP and probe/PEG6k-GNP is explained by the more efficient access of target DNA to the probe DNA on GNPs (nucleation step) in comparison with probe/diluent-GNP and probe/PEG10k-GNP. This is due to (i) strong electrostatic repulsion between the target DNA and probe/diluent-GNP and (ii) the probe DNA (7.0 nm of length) on probe/PEG2k-GNP and probe/PEG6k-GNP protrudes from the PEG2k layer (3.0 nm of length) and PEG6k layer (4.5 nm of length). Note that the values of appkh, and appKeq for probe/PEG6k-GNP were similar to those of probe/PEG2k-GNP, even though the probe/PEG6k-GNP has a larger PEG molecular weight and a shorter protruding structure of probe DNA than probe/PEG2k-GNP. This indicates that the short protruding structure of probe DNA from PEG layers is enough to facilitate the formation of a nucleus for DNA hybridization on GNPs (Fig. 3b).
In conclusion, this study described the quantitative determination and comparison of the parameters (HE, appkh, and appKeq) for DNA hybridization on DNA-modified GNPs with different nano-interfaces. The probe/diluent-GNP with low probe DNA density showed slightly more efficient DNA hybridization than that of probe-GNP with high probe DNA density, even though the total number of DNA strands per particle (particle charge) on these two DNA-modified GNPs is the same. When nonionic PEGs instead of anionic diluent DNA were immobilized on GNPs, probe/PEG10k-GNP showed a higher HE and appkh value compared with the corresponding values of probe/diluent-GNP due to diminished electrostatic repulsion. A further increase in appkh values was observed for both probe/PEG2k-GNP and probe/PEG6k-GNP due to the presence of protruding structures of probe DNA from the PEG layers, facilitating the nucleation step. Comparison of probe/PEG2k-GNP and probe/PEG6k-GNP showed that DNA hybridization on GNPs is facilitated even by a short protrusion of probe DNA from the PEG layers. The probe DNA density (particle charges) and protruding structure of probe DNA from the PEG layer were found to be the key factors for DNA hybridization on GNPs. Therefore, control of these factors is important for rational design of DNA-modified GNP-based sensing systems. However, further study using appropriate GNP samples seems to be necessary to clarify the DNA hybridization mechanism including nucleation step and zipping step, and we would like to report this issue elsewhere.
Acknowledgements
This work was partially supported by a Grant-in-Aid for Scientific Research (C) (no. 24510165 and no. 15K04632) from the Japan Society for the Promotion of Science (JSPS) and Mitsui Sumitomo Insurance Welfare Foundation.Notes and references
- C. A. Mirkin, R. L. Letsinger, R. C. Mucic and J. J. Storhoff, Nature, 1996, 382, 607–609 CrossRef CAS PubMed.
- K. C. Saha, S. S. Agasti, C. Kim, X. Li and V. M. Rotello, Chem. Rev., 2012, 112, 2739–2779 CrossRef CAS PubMed.
- Y.-W. Lin, C.-C. Huang and H.-T. Chang, Analyst, 2011, 136, 863–871 RSC.
- D. Kato and M. Oishi, ACS Nano, 2014, 8, 9988–9997 CrossRef CAS PubMed.
- A. Heuer-Jungemann, R. Kirkwood, A. H. El-Sagheer, T. Brown and A. G. Kanaras, Nanoscale, 2013, 5, 7209–7212 RSC.
- M. Oishi, S. Nakao and D. Kato, Chem. Commun., 2014, 50, 991–993 RSC.
- M. Oishi, Anal. Bioanal. Chem., 2015, 407, 4165–4172 CrossRef CAS PubMed.
- P. S. Randeria, M. R. Jones, K. L. Kohlstedt, R. J. Banga, M. Olvera de la Cruz, G. C. Schatz and C. A. Mirkin, J. Am. Chem. Soc., 2015, 137, 3486–3489 CrossRef CAS PubMed.
- A. K. R. Lytton-Jean and C. A. Mirkin, J. Am. Chem. Soc., 2005, 127, 12754–12755 CrossRef CAS PubMed.
- J. Xu and S. L. Craig, J. Am. Chem. Soc., 2005, 127, 13227–13231 CrossRef CAS PubMed.
- C. Chen, W. Wang, J. Ge and X. S. Zhao, Nucleic Acids Res., 2009, 37, 3756–3765 CrossRef CAS PubMed.
- H. Pei, F. Li, Y. Wan, M. Wei, H. Liu, Y. Su, N. Chen, Q. Huang and C. Fan, J. Am. Chem. Soc., 2012, 134, 11876–11879 CrossRef CAS PubMed.
- J.-H. Oh and J.-S. Lee, Anal. Chem., 2011, 83, 7364–7370 CrossRef CAS PubMed.
- V. A. Bloomfield, D. M. Crothers and I. Tinoco, Nucleic Acids: Structures, Properties, and Functions, University Science Books, Sausalito, 2000 Search PubMed.
- K. Rahme, L. Chen, R. G. Hobbs, M. A. Morris, C. O'Driscoll and J. D. Holmes, RSC Adv., 2013, 3, 6085–6094 RSC.
- L. M. Demers, C. A. Mirkin, R. C. Mucic, R. A. Reynolds, R. L. Letsinger, R. Elghanian and G. Viswanadham, Anal. Chem., 2000, 72, 5535–5541 CrossRef CAS.
- A. Halperin, A. Buhot and E. B. Zhulina, Biophys. J., 2005, 89, 796–811 CrossRef CAS PubMed.
- A. W. Peterson, R. J. Heaton and R. M. Georgiadis, Nucleic Acids Res., 2001, 29, 5163–5168 CrossRef CAS PubMed.
- M. M. A. Sekar, W. Bloch and P. M. St John, Nucleic Acids Res., 2005, 33, 366–375 CrossRef CAS PubMed.
- A. W. Peterson, L. K. Wolf and R. M. Georgiadis, J. Am. Chem. Soc., 2002, 124, 14601–14607 CrossRef CAS PubMed.
- P. V. Riccelli, F. Merante, K. T. Leung, S. Bortolin, R. L. Zastawny, R. Janeczko and A. S. Benight, Nucleic Acids Res., 2001, 29, 996–1004 CrossRef CAS PubMed.
- T. Liebermann, W. Knoll, P. Sluka and R. Herrmann, Colloids Surf., A, 2000, 169, 337–350 CrossRef CAS.
- M. R. Henry, P. W. Stevens, J. Sun and D. M. Kelso, Anal. Biochem., 1999, 276, 204–214 CrossRef CAS PubMed.
- K. K. Jensen, H. Ørum, P. E. Nielsen and B. Nordén, Biochemistry, 1997, 36, 5072–5077 CrossRef CAS PubMed.
- D. Porschke and M. Eigen, J. Mol. Biol., 1971, 62, 361–381 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra13116b |
| This journal is © The Royal Society of Chemistry 2015 |