
Umbelliprenin and lariciresinol isolated from a long-term-used herb medicine Ferula sinkiangensis induce apoptosis and G0/G1 arresting in gastric cancer cells†
Abstract
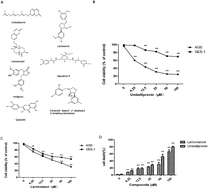
Effective chemicals isolated from folk medicine are commonly used in the treatment of cancer in Asian countries like China and India. Ferula sinkiangensis K. M. Shen is a traditional herb medicine used for treating stomach disorders in Xinjiang District of China for thousands of years. Here, we showed that the growth inhibition effects of seven compounds first isolated from the seeds of this herb in human gastric cancer cells and human normal gastric epithelium cells. Furthermore, we characterized the mechanism of the antiproliferation effects on gastric cancer cells of the two most specific and effective compounds: umbelliprenin (UM) and lariciresinol (LA). Annexin V/PI staining demonstrated that UM and LA induce apoptosis in gastric cancer AGS cells. Loss of mitochondrial membrane potential, upregulation of proapoptotic protein BAX, and activation of caspase 3 and PARP suggested that UM and LA caused the activation of the mitochondrial apoptosis pathway. Cell cycle analysis showed that UM and LA arrest cell cycle at G0/G1 phase. Western blot results showed that the expression of P53, P27, P16 and Rb proteins increased, while the expression of cyclin D, cyclin E, Cdk4 and Cdk2 decreased in cancer cells. Overall, these data provided evidence that UM and LA have the potential to be used in cancer therapy.
 Please wait while we load your content...
Please wait while we load your content...

