
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Modulation of the stability of i-motif structures using an acyclic threoninol cytidine derivative†
Sonia
Pérez-Rentero
 a,
Raimundo
Gargallo
b,
Carlos
González
c and
Ramon
Eritja
*a
a,
Raimundo
Gargallo
b,
Carlos
González
c and
Ramon
Eritja
*a
aInstitute for Advanced Chemistry of Catalonia (IQAC-CSIC), CIBER-BBN, Jordi Girona 18-26, E-08034 Barcelona, Spain. E-mail: recgma@cid.csic.es
bDepartment of Analytical Chemistry, University of Barcelona, Martí i Franqués 1-11, E-08028 Barcelona, Spain
cInstitute of Physical Chemistry “Rocasolano”, CSIC, Serrano 119, E-28006 Madrid, Spain
First published on 20th July 2015
Abstract
The effect of aTNA (acyclic threoninol nucleic acids) units on the stability of intramolecular i-motifs was investigated by spectroscopic techniques. The replacement of selected positions of the C-core can modulate the stability at different pH ranges.
The i-motif is a secondary structure found in C-rich DNA sequences, which is stabilised under acidic conditions. The structure comprises two strands arranged in a parallel-type duplex with hemiprotonated C·CH+ base pairs, and two of these duplexes are held together in an antiparallel fashion by intercalation of each C·CH+ base pair (Fig. 1a).1
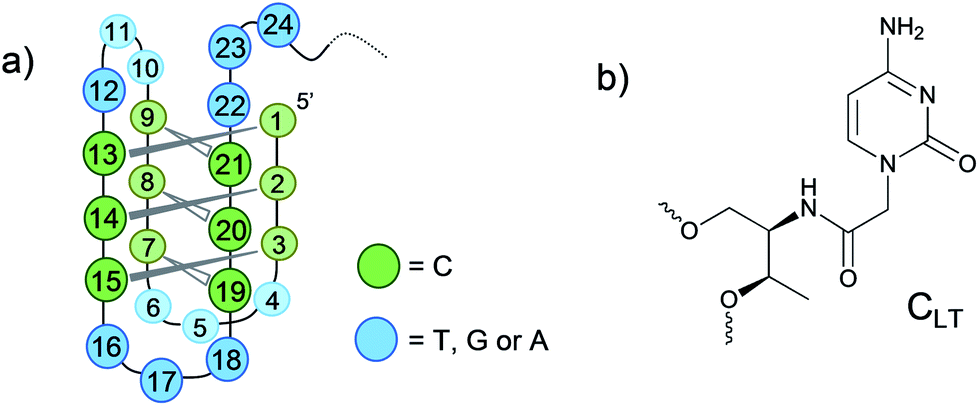 | ||
| Fig. 1 (a) Schematic representation of a potential folding topology of the intermolecular i-motif studied in this work. (b) Chemical structure of CLT. | ||
From the biological point of view, the presence of i-motif-forming sequences in telomeric and promoter regions supports the hypothesis that they may be functionally relevant.2 Recently, the role of these structures as modulators of gene expression has been assessed.3 Additionally, these structures are interesting in the field of nanotechnology, since for instance they have been used to control the assembly of supramolecular structures4 or in the design of molecular motors5 and sensing systems such as nanoswitches driven by pH changes to monitor biological processes.6 Desirable properties regarding this last application points towards the necessity of nuclease resistant i-motifs and the availability of different pH-sensors suited to different pH regimes.6a The stability of i-motif structures is highly dependent of the base sequence.7 It has been reported that simple manipulations of the C-core8 or the loops9 can modulate the pH sensitivity and also the introduction of chemical modifications.2 In particular, sugar modifications of the cytidine core4 have pointed the relevance of the sugar–sugar contacts in the stability of i-motifs.10
Threoninol is an acyclic scaffold that has been used to design new artificial nucleic acids (aTNA), capable of forming exceptionally stable homo-duplexes.11
Moreover, aTNA's can form stable hetero-duplexes with DNA and RNA in a parallel manner,12 and also they are compatible with parallel G-quadruplex structures.13 Recently, this acyclic scaffold has been used to design functional RNA/aTNA chimeras with improved nuclease resistance.14 To date the effect of this modification in the formation of i-motifs has not been tested. In sight of these results, we are interested in study the effect of this sugar replacement on the thermal and pH stability.
The present work is based on a well described C-rich fragment of the vertebrate telomere (C3TA2)3C3T, which folds intramolecularly, where four cytidine stretches connected by three TAA loops form six C·CH+ base pairs.15 In this work the loop bases were substituted by thymines and then, different positions of the C-core stretches of the 24 nt fragment (C3T3)4(Lt_0) were systematically replaced by CLT units (Fig. 1b), (Table 1). For detailed information see ESI† (Sections 6–8). Lt_0 was modified by a single CLT substitution at various positions of the 5′-terminal stretch (Lt_1, Lt_2 and Lt_3) and more internal positions (Lt_13, Lt_14 and Lt_15). Additionally, oligonucleotides containing two CLT modifications involved in the same C·CH+ base pair (Lt_1/13, Lt_2/14 and Lt_3/15) were studied. Finally, Lt_1/2/3 with three consecutive C modifications in the 5′-terminal stretch was included in the analysis. We assessed the thermodynamic stability of the modified 24 nt fragments by monitoring the absorbance at 295 nm as a function of temperature (Fig. S1, ESI†) and compared with the non-modified version (Table 1). For additional information see ESI† (Section 2).
| Nameb | T m | ΔTm | ΔG0298 | ΔΔG0298 | ΔH0 | TΔS0298 |
|---|---|---|---|---|---|---|
| a T m is expressed in °C and ΔG0298, ΔH0 and TΔS0298 in kcal mol−1. Experimental errors are shown in Table S1 (see ESI). b The numbers indicate the positions substituted. Coligonucleotide = 2.7 μM. | ||||||
| Lt_0 | 47.8 | — | −5.7 | — | −79.3 | −73.7 |
| Lt_1 | 39.5 | −8.3 | −2.9 | 2.8 | −61.7 | −58.8 |
| Lt_2 | 29.1 | −18.7 | −0.6 | 5.1 | −45.6 | −45.0 |
| Lt_3 | 38.4 | −9.4 | −2.6 | 3.1 | −60.9 | −58.4 |
| Lt_13 | 37.4 | −10.4 | −2.7 | 3.0 | −67.4 | −64.8 |
| Lt_14 | 29.3 | −18.5 | −0.8 | 4.9 | −56.3 | −55.5 |
| Lt_15 | 37.9 | −9.9 | −2.7 | 3.0 | −65.1 | −62.4 |
| Lt_1/13 | 35.4 | −12.4 | −2.3 | 3.4 | −67.5 | −65.2 |
| Lt_2/14 | 32.4 | −15.4 | −1.6 | 4.1 | −66.4 | −64.8 |
| Lt_3/15 | 34.7 | −13.1 | −2.1 | 3.6 | −65.8 | −63.7 |
| Lt_1/2/3 | 35.6 | −12.2 | −2.4 | 3.3 | −69.3 | −67.0 |
At pH 5.3 the UV melting analyses reveal that all the CLT substitutions reduce significantly the Tm, and the effect is highly dependent of the position. Single CLT substitutions near the loops are less destabilizing than those in the middle of C-tracts. Similar effect has been observed with other acyclic backbone modifications.10d The effect of CLT substitutions is not additive. When the two cytidines involved in a C·CH+ base pair were replaced, the Tm is slightly lower than in single substitutions. The effect is opposite when the C·CH+ is located in the center of the C-tracts (Lt_2/14). Finally, Lt_1/2/3 shows a higher thermal stability than some of the oligonucleotides containing one or two modifications. In all cases the values of Gibb's free energies follow the same pattern observed with the Tm values as expected because depend on the equilibrium constant. Analysing the enthalpic term for oligonucleotides containing one or two CLT substitutions near the loops, it can be observed than the ΔH0 exhibit a similar decrease indicating than the destabilization is mostly of enthalpic origin. The influence of the enthalpic change is compensated by opposite change in the entropy term leaving the influence on Gibbs's energy rather modest. A single replacement in central positions of a cytidine core causes a higher drop of ΔH0, especially when is implicated the 5′-terminal stretch, suggesting that the C·CH+ base pair is affected and additionally the distortion may be affecting the neighbouring base pairing. Surprisingly, when both modifications occupy the central part of the core, it leads to a more stable structure than the ones obtained with Lt_2 and Lt_14. Taking into account that in this case the enthalpic term is more favourable, it can be reasoned that the pairing dC–CLT is less tolerated than the CLT–CLT pair. Finally, Lt_1/2/3 has the highest enthalpic term, but at the same time the most unfavourable entropic term.
Also we investigated the i-motif formation by CD spectroscopy. In Fig. 2 there are depicted a selection of some CD spectra. For detailed information see ESI (Fig. S2 and Table S2†). Lt_0 has the characteristic i-motif signature,16 with positive and negative peaks around 287 and 262 nm respectively and as well all modified sequences show this specific profile. Comparing with Lt_0, in general the modifications produce blue shifts of the positive and/or negative bands and a decrease in the molar ellipticity to a greater or lesser extent. The most affected is the version containing three CLT residues, followed by the oligonucleotides containing a substitution in an internal position of the 5′-terminal stretch. As CD spectroscopy is sensitive to structural changes,17 it can be reasoned that the deviations in the CD spectra from the natural sequence would be more pronounced as higher is the degree of distortion introduced in the i-motif structure.
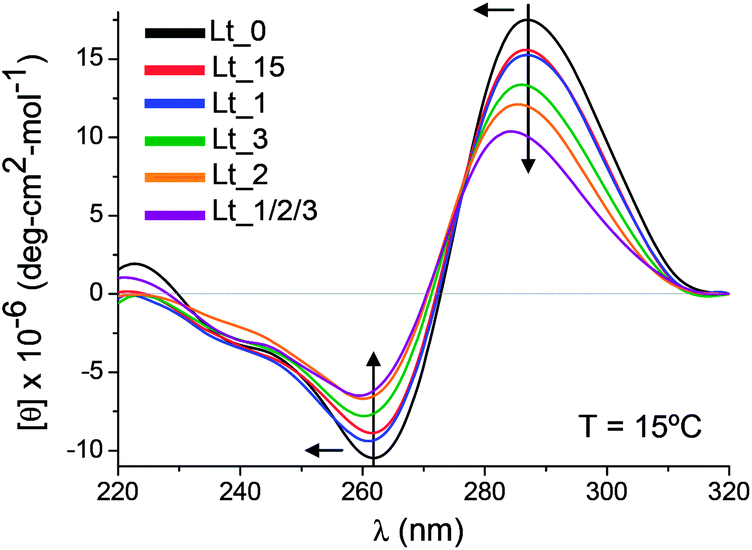 | ||
| Fig. 2 CD spectra of Lt_0 and modified oligonucleotides at pH 5.3. | ||
Since structural changes may affect the pH stability, we decided to compare the pH dependence of two modified oligonucleotides with the unmodified version. We selected the one showing the most altered CD spectrum (Lt_1/2/3) and one showing moderate changes (Lt_3). The unfolding of the i-motif with pH at 25 °C was monitored at 295 nm. By plotting the unfolded fraction against pH and adjusting to a sigmoidal (see Section 4 ESI†) we obtained the titration curves showed in Fig. 3. The pH transition midpoint (pHT) and the transitional range (RT, range between 10 and 90% of unfolded structure) were calculated from the adjusted curves (Table 2). Comparing Lt_3 with Lt_0, the RT has been slightly widened and also shifted toward acidic pH values, showing a pHT around pH 6.00. Lt_1/2/3 as well shows a decrease in the pHT, but the RT is almost 3-fold wider. The presence of three consecutive CLT in the 5′-external stretch introduces higher flexibility.
 | ||
| Fig. 3 pH titration of Lt_0, Lt_3 and Lt_1/2/3. | ||
| Name | pHT | pH10 | pH90 | R T |
|---|---|---|---|---|
| Lt_0 | 6.54 ± 0.02 | 6.05 ± 0.02 | 6.71 ± 0.02 | 0.66 ± 0.04 |
| Lt_3 | 6.00 ± 0.08 | 5.42 ± 0.02 | 6.20 ± 0.07 | 0.78 ± 0.09 |
| Lt_1/2/3 | 6.16 ± 0.02 | 4.80 ± 0.02 | 6.70 ± 0.02 | 1.90 ± 0.04 |
These effects may lead to the formation of several misfolded conformations provoking a decreased cooperativity.
Next, we performed 1D 1H NMR experiments with Lt_0, Lt_3 and Lt_1/2/3 at different temperatures to obtain more detailed structural information (see Fig. S4, ESI†). Fig. 4 shows for each oligonucleotide the experiments recorded at 5 °C. In the three cases, the NMR spectra are consistent with i-motif formation. The characteristic imino signals of protonated cytosines are clearly observed at 15.0–16.0 ppm,7,15 as well as the thymine imino protons in the loop regions (around 11.0 ppm), in agreement with previous studies reported in the literature.15 The differences in the exchangeable protons spectra between the unmodified and Lt_3 and Lt1/2/3 are significant, indicating the effect of the acyclic threoninol backbone propagates up to the base pairs, affecting the core of the i-motif structure. Such effect is particularly pronounced in the case of Lt/1/2/3. The unfolding of the structure with increasing temperature can be followed in Fig. S4 in ESI.† In the unmodified oligonucleotide, the imino protons are observed up to 45 °C. In agreement with UV melting experiments, the exchangeable protons signals of Lt_3 and Lt_1/2/3 disappear at lower temperatures than the unmodified oligonucleotide.
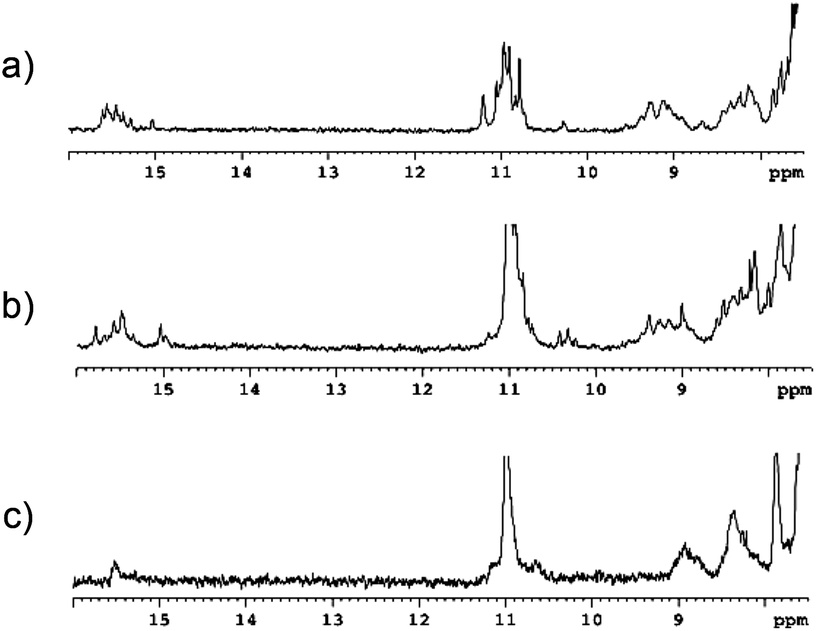 | ||
| Fig. 4 NMR spectra at 5 °C and pH = 4.2 of (a) Lt_0, (b) Lt_3 and (c) Lt_1/2/3. | ||
In summary, we evaluated for the first time the effect of deoxy-sugar replacements by CLT substitutions in the ability to form intramolecular i-motifs. Different positions of the C-core were replaced and the thermal and thermodynamic stability have been studied. In general the CLT modifications reduced significantly the stability of the resulting structures and the effect is dependent of the position and the type of pair implicated, being the pairing dC–CLT less favourable than the CLT–CLT pair. When a single substitution was introduced, the positions with a lesser decrease in stability are the positions near the loops.
In addition the pH stability of Lt_3 and Lt_1/2/3 was studied. In comparison with Lt_0, a single CLT substitution produced a decrease of pHT of about 0.54 units and maintained a comparable range of response. The substitution of three consecutive cytidines led to a less cooperative transition. Recently it has been described than the L-threoninol nucleic acids are more stable to nuclease degradation than unmodified oligonucleotides.14 For this reason the possibility of modulating the pH stability of i-motifs by selected replacements of cytidines by CLT modifications may allow the design of nuclease resistant i-motif structures suited for different pH ranges.
Acknowledgements
This work was supported by Spanish Ministry of Economy and Competitiveness (grants CTQ2014-52588-R and CTQ2012-38616-C02-02), the Generalitat de Catalunya (grant 2014 SGR 1106; 2014 SGR 187) and the Communities MULTIFUN (contract NMP4-LA-2011-262943). CIBER-BBN is an initiative funded by the VI National R&D&I Plan 2008–2011, Iniciativa Ingenio 2010, Consolider Program, CIBER Actions and financed by the Instituto de Salud Carlos III with assistance from the European Regional Development Fund.References
- K. Gehring, J. L. Leroy and M. Guéron, Nature, 1993, 363, 561–565 CrossRef CAS PubMed.
- (a) S. Benabou, A. Aviñó, R. Eritja, C. González and R. Gargallo, RSC Adv., 2014, 4, 26956–26980 RSC; (b) H. A. Day, P. Pavlou and Z. A. E. Waller, Bioorg. Med. Chem., 2014, 22, 4407–4418 CrossRef CAS PubMed.
- H.-J. Kang, S. Kendrick, S. M. Hecht and L. H. Hurley, J. Am. Chem. Soc., 2014, 136, 4172–4185 CrossRef PubMed.
- (a) H. B. Ghoedke, R. Krishnan, K. Vignesh, G. Kumar, C. Narayana and Y. Krishnan, Angew. Chem., Int. Ed., 2007, 119, 2700–2703 CrossRef; (b) E. Guittet, D. Renciuk and J.-L. Leroy, Nucleic Acids Res., 2012, 40(11), 5162–5170 CrossRef CAS PubMed.
- D. Liu and S. Balasubramanian, Angew. Chem., Int. Ed., 2003, 42, 5734–5736 CrossRef CAS PubMed.
- (a) S. Modi, C. Nizak, S. Surana, S. Halder and Y. Krishnan, Nat. Nanotechnol., 2013, 8, 459–467 CrossRef CAS PubMed; (b) S. Surana, J. M. Bhat, S. P. Koushika and Y. Krishnan, Nat. Commun., 2011, 2, 339–345 CrossRef PubMed.
- J.-L. Mergny, L. Lacroix, X. Han, J.-L. Leroy and C. Hélène, J. Am. Chem. Soc., 1995, 6, 8887–8898 CrossRef.
- I. V. Nesterova and E. E. Nesterov, J. Am. Chem. Soc., 2014, 136, 8843–8846 CrossRef CAS PubMed.
- S. P. Gurung, C. Schwarz, J. P. Hall, C. J. Cardin and J. A. Brazier, Chem. Commun., 2015, 51, 5630–5632 RSC.
- (a) C. P. Fenna, V. J. Wilkinson, J. R. P. Arnold and R. Cosstick, Chem. Commun., 2008, 3567–3569 RSC; (b) N. Kumar, M. Petersen and S. Maiti, Chem. Commun., 2009, 1532–1534 RSC; (c) S. Modi, A. H. Wani and Y. Krishnan, Nucleic Acids Res., 2006, 34(16), 4354–4363 CrossRef CAS PubMed; (d) A. Pasternak and J. Wengel, Bioorg. Med. Chem. Lett., 2011, 21, 752–755 CrossRef CAS PubMed.
- H. Asanuma, T. Toda, K. Murayama, X. Liang and H. Kashida, J. Am. Chem. Soc., 2010, 132, 14702–14703 CrossRef CAS PubMed.
- K. Murayama, H. Kashida and H. Asanuma, Chem. Commun., 2015, 51, 6500–6503 RSC.
- J. Zhou, K. Murayama, S. Amrane, F. Rosu, H. Kashida, A. Bourdoncle, H. Asanuma and J.-L. Mergny, Chem. Sci., 2013, 4, 3693–3698 RSC.
- (a) A. Alagia, M. Terrazas and R. Eritja, Molecules, 2014, 19, 17872–17896 CrossRef CAS PubMed; (b) A. Alagia, M. Terrazas and R. Eritja, Molecules, 2015, 20, 7602–7619 CrossRef CAS PubMed.
- A. T. Phan, M. Guéron and J.-L. Leroy, J. Mol. Biol., 2000, 299, 123–144 CrossRef CAS PubMed.
- G. Manzini, N. Yathindra and L. E. Xodo, Nucleic Acids Res., 1994, 22(22), 4634–4640 CrossRef CAS PubMed.
- J. Kypr, I. Kejnovská, D. Renčiuk and M. Vorlíčková, Nucleic Acids Res., 2009, 37(6), 1713–1725 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Experimental methods, synthesis and characterizations of CLT monomers and oligonucleotides. See DOI: 10.1039/c5ra10096h |
| This journal is © The Royal Society of Chemistry 2015 |