
Time-dependent antioxidative responses of ramie (Boehmeria nivea (L.) Gaudich) to moderate cadmium stress and its up-regulation mechanism by spermidine antioxidant
Lu Zhouab,
Yunguo Liu*ab,
Xi Huc,
Guangming Zengab,
Yaqin Wangab,
Xinjiang Huab,
Yaoyu Zhouab,
Xiaofei Tanab,
Luhua Jiangab and
Xiaoxia Zengab
aCollege of Environmental Science and Engineering, Hunan University, Changsha 410082, P.R. China. E-mail: yunguoliu_hnu@163.com; Fax: +86 731 88823701; Tel: +86 731 88649208
bKey Laboratory of Environmental Biology and Pollution Control (Hunan University), Ministry of Education, Changsha 410082, P.R. China
cCollege of Environmental Science and Engineering Research, Central South University of Forestry and Technology, Changsha 410004, P.R. China
First published on 3rd September 2015
Abstract
Cadmium (Cd) is present in soil and water at low concentrations that can be harmful to the biota. A Cd-tolerant plant species named ramie (Boehmeria nivea (L.) Gaudich) was applied to probe the precise Cd tolerance mechanism with different treatment duration and the up-regulation effect for antioxidant capacity by spermidine (Spd) pretreatment. The results showed that short-term (0–7 d) Cd stress resulted in the enhancement of pigment content, the accumulation of soluble sugar and the activation of partial antioxidants, meanwhile reducing the level of malondialdehyde (MDA), which is the indicator of oxidative stress. Spd pretreatment markedly increased the soluble protein, soluble sugar and reduced glutathione (GSH) content in ramie leaves under short-term Cd stress, whereas, it showed no significantly beneficial effects on photosynthesis and other antioxidants. Long-term (0–15 d) Cd stress may lead to growth inhibition which is associated with Cd accumulation, protein oxidation, sugar degradation, MDA surge and antioxidant inactivation. Spd application markedly alleviated long-term Cd toxicity by decreasing Cd accumulation, stabilizing cellular macromolecules such as pigments, soluble protein and sugar and inhibiting peroxidase (POD), catalase (CAT) and malate dehydrogenase (MDH) inactivation and GSH degradation. Our present work concluded that ramie could tolerate moderate Cd stress during a certain time span. Spd application could play a vital role in the promotion of Cd tolerance by adjusting different kinds of parameters.
1. Introduction
Heavy metals are generally present at low concentration in soil and water.1 Among the heavy metals, cadmium (Cd) is of special concern because of its high toxicity and large solubility, mainly derived from combustion of fossil fuels, unreasonable fertilizer application, mining and other industrial processes.2 As a non-essential trace element, Cd can be taken up readily by plant roots and transported to the vegetative and reproductive organs, where it shows phytotoxicity even at low doses.3 Toxic effects of Cd are manifested by various symptoms of injury, including chlorosis, necrosis, leaf rolling, wilting and growth inhibition.2,4It is well-known that oxidative stress is a rapid response to excessive Cd exposure, which is reflected in the abnormal upsurge of reactive oxygen species (ROS), such as superoxide (O2˙−), hydrogen peroxide (H2O2) and hydroxyl radical (˙OH).2 Although ROS may function as signaling molecules involved in mediating responses to heavy metal stress at trace concentration, they can be extremely harmful to organisms by oxidizing proteins, chlorophyll, lipids, nucleic acids and carbohydrates if the stress becomes high and/or continues for a prolonged period.5 Malondialdehyde (MDA), a cytotoxic product of lipid peroxidation, is generally used as an indicator of oxidative stress due to plant membrane is one of the first structures to suffer from the ROS attack.6 Oxidative stress, however, is essentially a regulated process. The proliferation of ROS is tightly controlled by versatile and cooperative antioxidative defense mechanisms, which maintain the balance between ROS production and decomposition and the normal redox homeostasis in cells. One of the defense mechanisms is the activation of various enzymatic antioxidants such as superoxide (SOD), peroxidase (POD), catalase (CAT), malate dehydrogenase (MDH), etc.7 The other is the accumulation or depletion of non-enzymatic metabolites including glutathione, ascorbic acid, proline, polyamine, α-tocopherols and carotenoid. Reduced glutathione (GSH), a major non-enzymatic metabolite abundant in plant cells, is considered as the most crucial intracellular defense against oxidative stress through controlling the cellular redox state and ROS accumulation.4,8 Antioxidative defense mechanisms may provide a strategy to enhance plant oxidative stress tolerance. However, under severe stress conditions, the antioxidant capacity may not be sufficient to attenuate the harmful effects of oxidative injury with the balance of ROS shifting, and then resulting in membrane damage, inactivation of enzymes, pigment loss and the metabolic disturbance of essential elements.4,8
Polyamines (PAs), phytohormone-like aliphatic nitrogenous compounds, are involved in both plant physiological processes and oxidative stress tolerance. The most abundantly found PAs in plants are putrescine (Put), spermidine (Spd) and spermine (Spm).9 It has been proposed that exogenously applied PAs especially the triamine Spd can act as an antioxidant in intracellular metabolic processes, thus ameliorating metal-induced oxidative damage.10,11 However, multiplicity in the literature exists in the metal detoxification mechanisms of PAs. Firstly, PAs are able to be bond to negatively charged proteins, nucleic acids, membrane phospholipids and cell wall constituents owing to their polycationic nature at physiological pH, resulting in the stabilization of cell membrane and inhibition of lipid peroxidation.12 Secondly, PAs may either act directly as oxygen radical scavengers or form a higher potential of antioxidant capacity by adjusting enzyme activity. Moreover, pretreated PAs may serve as a stress signal to activate acclimation and defense mechanisms, thereby effectively resisting the following heavy metal stress.12,13 Despite extensive studies on PAs, it is essential to obtain more precise information on the timing and location of PAs actions for antioxidative defense response in plants due to its high price.
Ramie (Boehmeria nivea (L.) Gaudich), a textile crop that extensively grown in China, was selected as the test plant due to its high biomass, Cd tolerance and economic value.14 In our earlier studies, we have reported the uptake and distribution pattern of Cd in ramie and found that growth inhibition was a crucial factor limiting application of ramie in phytoremediation in contaminated sites. The co-workers have been attempting to improve the Cd tolerance of ramie by various exogenous substances such as nitric oxide, selenium, citric and oxalic acids.15–17 Recently, PAs have received attention in the framework of phytoremediation intending to decontaminate polluted soil, water and mine tailing with high biomass plants.18,19 However, the mechanism of Cd tolerance in ramie seedlings in combination with PAs has rarely been reported. In this work, hydroponic experiments were conducted to investigate the effect of moderate level Cd on the tolerance mechanism in ramie seedlings with different treatment duration and the role of Spd in promoting antioxidative defense mechanisms to cope with Cd stress. The main aims of this study were to evaluate whether exogenous Spd enhanced short-term or long-term Cd tolerance with reference to: the changes in (1) Cd accumulation; (2) cellular macromolecules metabolism; (3) membrane lipid peroxidation and (4) antioxidative capacity.
2. Materials and methods
2.1. Plant materials and cultivation conditions
One-month-old ramie seedlings were originally collected from the Ramie Institute of Hunan Agricultural University in China. Plants were washed with distilled water and acclimatized in a hydroponic system with 10% Hoagland nutrient solution for two weeks. Afterwards, plants of similar height and weight were selected for experimentation and grown in a growth chamber with 25 °C/20 °C (day/night) temperature, 12 h photoperiod, light intensity PAR 300 μmol m−2 s−1 and 60 ± 5% relative humidity.2.2. Treatments in hydroponic experiment
For Spd pretreatment, plants were sprayed with 0.1 mM Spd (1 mL per plant) at 12 h before the application of Cd, while controls were sprayed with distilled water. Tween 80 solution (0.2% v/v) was used as a surfactant. Then Cd treatments in triplicates were as follows: (1) controls in 10% Hoagland solution (CK); (2) 10% Hoagland solution + 0.1 mM Spd (Spd); (3) 10% Hoagland solution + 30 μM Cd (Cd); (4) 10% Hoagland solution + 30 μM Cd + 0.1 mM Spd (Cd + Spd). The Cd concentration was calculated on Cd2+ ion and equivalent to 3.37 mg L−1. All treatment solutions were renewed every 3 d. The spraying of Spd and distilled water was arranged at 8:00 am and 20:00 pm of each day in the period of Cd treatment. After the indicated time intervals (0, 1, 3, 5, 7, 10 and 15 d), leaf segments from identical samples were collected and thoroughly washed for analysis. Cd and Spd concentrations used for treatments were selected according to the preliminary experimental results and the related literatures.20,212.3. Quantification of Cd accumulation and dry biomass
After 15 d of Cd treatments, plant roots were rinsed thoroughly with 0.1 M HNO3. Plant tissues were separated, oven dried at 80 °C for 2 d to a constant weight, then obtained for the determination of dry biomass. Powdered dry samples (0.5 g) were digested with 4 mL of HNO3/HClO4 (3![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1, v/v), then dissolved in the 10 mL of 0.1 M HNO3. The Cd contents were then measured by atomic absorption spectrophotometer (TAS-990F, Beijing Persee, China).
:1, v/v), then dissolved in the 10 mL of 0.1 M HNO3. The Cd contents were then measured by atomic absorption spectrophotometer (TAS-990F, Beijing Persee, China).
2.4. Determination of pigments content
For chlorophyll and carotenoid determinations, the leaves were homogenized in 10 mL of 95% ethanol in darkness and centrifuged at 4000g for 15 min. Then the absorbance of the supernatant was measured at 450, 645 and 663 nm with ultraviolet spectrophotometer (UV2550, Shimadzu, Japan), as described by Knudson.222.5. Determination of membrane lipid peroxide
MDA content represents the degree of leaf membrane lipid peroxidation.6 The content of MDA was measured by the thiobarbituric acid (TBA) reaction method with modification.23 Fresh leaves (0.5 g) were homogenized in a pre-chilled mortar and pestle with 10 mL of 10% (w/v) trichloroacetic acid (TCA) and then centrifuged at 10000g for 15 min. The mixture containing 1 mL aliquot of supernatant and 2 mL of 0.67% (w/v) TBA in 10% (w/v) TCA was heated to 100 °C for 15 min and rapidly cooled in an ice-bath. After centrifugation at 4 °C, the absorbance of supernatant was measured at 450, 532 and 600 nm, respectively (extinction coefficient was 155 mM−1 cm−1).
2.6. Determination of soluble protein and sugar
The soluble protein content was determined by spectrophotometric method of Bradford using bovine serum albumin (BSA) as standard.24 Leaves were ground with 3 mL extracting solution containing 50 mM Tris–HCl (pH 7.8), 0.5 mM MgCl2, 1 mM EDTA and 1 mM dithiothreitol in pre-chilled mortars. After centrifugation at 10000g at 4 °C for 20 min, the supernatant was mixed with 0.9 mL Tris buffer solution and 5 mL coomassie brilliant blue G-250 staining solution. The supernatant absorbance was measured at 595 nm.
The soluble sugar was assayed by the anthrone-sulfuric acid method, and its content was calculated using glucose as a standard.25 After added with 10 mL of distilled water, fresh leaves (0.2 g) were incubated in boiling water bath for 30 min, the supernatants were used for the determination of soluble sugar.
2.7. Assay of enzyme activities
All enzymatic extractions were carried out at 4 °C. Fresh leaves (0.2 g) were homogenized using 1.8 mL of 50 mM pre-chilled phosphate buffer (pH 7.8) containing 1% (w/v) polyvinylpyrrolidone (PVP) for SOD and POD activity detection, and 0.9% (w/v) NaCl for CAT and MDH. After centrifugation at 14000g for 20 min, the supernatant was extracted for assays of enzyme activity. The enzyme activities were determined using ELISA kits purchased from Jiancheng Biotech, Nanjing, China, strictly following the manufacturer's instructions. These enzyme activities were expressed as U g−1 fw.
2.8. Determination of reduced glutathione
As an effective non-enzymatic antioxidant, the content of glutathione was determined. Fresh leaves (0.2 g) were homogenized in pre-chilled mortars and pestles in TCA. Then the homogenates were centrifuged at 10000g for 10 min at 4 °C. The supernatants were used for assays of GSH content according to Ellman.26 The reaction solution contained NaH2PO4 (2 mM, pH = 7.4), 5,50-dithio-bis(2-nitrobenzonic) (6 mM) and 0.2 mL of supernatant. After incubation at 30 °C for 5 min, the mixture absorbance was assayed at 412 nm.
2.9. Statistical analysis
The results are presented as means ± standard deviations (SD) of three replications. The different significance of short-term (0–7 d) or long-term (0–15 d) Cd treatment between control group and experimental group was analyzed by one-way analysis of variance (ANOVA) at the 95% confidence limits level, taking P < 0.05 as significantly different according to LSD test.3. Results
3.1. Cd accumulation and plant biomass
After 15 d of Cd treatment, the distribution of Cd in different organ and plant dry biomass are shown in Table 1. No matter whether with Spd pretreatment or not, Cd concentration in different organ decreased following the order of roots > shoots > leaves. Exogenous Spd application alleviated Cd accumulation in each organ, resulting in that the Cd concentration in root, shoot and leaf tissues decreased by 17%, 10% and 11%, respectively. Meanwhile, the application of Spd slightly improved the transfer factor, which suggested a promotion effect on Cd translocation from roots to shoots and leaves. For dry biomass, exogenous Spd pretreatment significantly alleviated the Cd-mediated reduction (P < 0.05).| Treatment | Cd concentration (mg kg−1 dw) | TF value (%) | Dry biomass (g per plant) | |||
|---|---|---|---|---|---|---|
| Root | Shoot | Leave | Root | Shoot | ||
| CK | ND | ND | ND | — | 4.28 ± 0.31 | 1.82 ± 0.12 |
| Spd | ND | ND | ND | — | 4.42 ± 0.27 | 2.04 ± 0.11 |
| Cd | 368.23 ± 35.74 | 33.17 ± 2.93 | 24.86 ± 2.25 | 15.76 | 2.91 ± 0.18 | 1.31 ± 0.05 |
| Cd + Spd | 305.42 ± 29.15 | 29.82 ± 2.47 | 22.19 ± 1.74 | 17.00 | 4.08 ± 0.26 | 1.73 ± 0.09 |
3.2. Photosynthetic pigment contents
A time-dependent response of Cd stress was observed on the photosynthetic pigments in ramie leaves (Fig. 1 and 2A). The pigment levels under the CK condition presented a stable tendency with the time prolongation, whereas Spd pretreatment resulted in significant enhancement of both chlorophyll and carotenoid in the CK levels (P < 0.05). With Cd treatment, both chlorophyll and carotenoid contents were significantly increased within 7 d (P < 0.05), and then declined below the CK level at 15 d. The chlorophyll content increased significantly in the Cd + Spd treatment group compared with Cd stress alone after 15 d (P < 0.05), but exhibited no obvious change during short-term Cd treatment. The ratio of chlorophyll-a (Chl-a) to chlorophyll-b (Chl-b) gradually dropped during long-term Cd exposure, whereas maintained steady with Spd application. | ||
| Fig. 1 Effects of cadmium (Cd) and spermidine (Spd) application on chlorophyll content in the leaves of ramie seedlings after different periods. Each data represents means ± SD of three replicates. Asterisks (*) indicate significant differences (P < 0.05) among the four treatments. CK, Spd, Cd and Cd + Spd correspond to 10% Hoagland solution (HS), HS + 0.1 mM Spd, HS + 30 μM Cd and HS + 30 μM Cd + 0.1 mM Spd, respectively. | ||
 | ||
| Fig. 2 Effects of cadmium (Cd) and spermidine (Spd) application on carotenoid (A), soluble protein (B) and soluble sugar (C) contents in the leaves of ramie seedlings after different periods. Each data represents means ± SD of three replicates. Asterisks (*) indicate significant differences (P < 0.05) among the four treatments. CK, Spd, Cd and Cd + Spd correspond to 10% Hoagland solution (HS), HS + 0.1 mM Spd, HS + 30 μM Cd and HS + 30 μM Cd + 0.1 mM Spd, respectively. | ||
3.3. Protein and sugar metabolism
As shown in Fig. 2B, Spd pretreatment caused great induction of soluble protein content in the leaves of CK plants (P < 0.05). After Cd treatment for 5 d, the soluble protein content showed no significant difference compared with the CK level. Continuous Cd toxicity caused a slight increase in soluble protein content and then exhibited a strong suppression in soluble protein synthesis. The soluble protein content reached its maximal after 10 d, about 1.4 times over the CK level. At the later stage, Spd pretreatment significantly alleviated the decrement of soluble protein content in ramie leaves by Cd stress (P < 0.05).The soluble sugar content, which is responsible for energy storage, is also shown in Fig. 2C. Cd treatment demonstrated considerably significant impact on the content of soluble sugar compared with the CK group (P < 0.05). The soluble sugar content was sharply increased since the 3rd day and it was 2.4 times of the CK level when Cd treatment time lasted for 10 d. After 10 d, however, the soluble sugar content showed downtrend in the Cd treatment group. Exogenous Spd could significantly promote the accumulation of soluble sugar through the entire Cd treatment (P < 0.05) and caused the greatest increase at the last day.
3.4. Lipid peroxidation
The effect of Spd pretreatment on MDA content of different test time is shown in Fig. 3. With the prolongation of Cd treatment time, the MDA content in ramie leaves showed a rising trend, though it went through a minimum after 7 d. Exogenous Spd was found to be very potent inhibitors of MDA formation during long-term Cd stress (P < 0.05) and the optimal time-point was at 15 d, at which the MDA content decreased by about 25%. For a short period of Cd treatment, however, pretreatment with Spd did not show significantly mitigative effect on lipid peroxidation caused by Cd stress.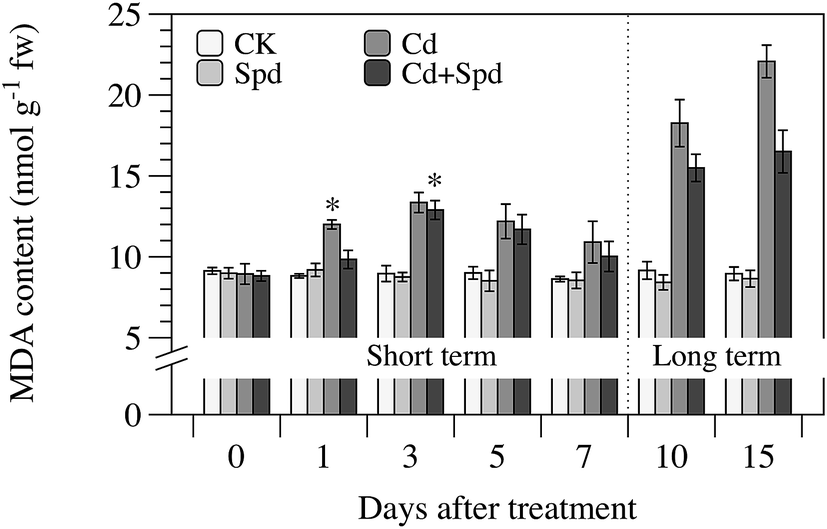 | ||
| Fig. 3 Effects of cadmium (Cd) and spermidine (Spd) application on malondialdehyde (MDA) content in the leaves of ramie seedlings after different periods. Each data represents means ± SD of three replicates. Asterisks (*) indicate significant differences (P < 0.05) among the four treatments. CK, Spd, Cd and Cd + Spd correspond to 10% Hoagland solution (HS), HS + 0.1 mM Spd, HS + 30 μM Cd and HS + 30 μM Cd + 0.1 mM Spd, respectively. | ||
3.5. Enzymatic antioxidants
Activities of several representative antioxidant enzymes, including SOD, POD, CAT and MDH were selected to evaluate the function of Spd in regulation of these antioxidant enzymes upon Cd stress. As can be seen from Fig. 4, overall, no significant changes of SOD, POD and CAT activity were found between the CK group and the Spd treatment group. Whether following short-term or long-term Cd toxicity, ramie leaves exhibited a remarkable elevation in the activity of SOD (P < 0.05), which reached the peak of 126.1 U g−1 fw at 10 d. Both POD and CAT activity, as opposed to SOD, showed no obvious increase within 7 d and then underwent downtrend caused by long-term Cd toxicity. After 15 d of Cd treatment, POD and CAT activity in the ramie seedlings reduced to 92% and 57% of the CK level, respectively. Under short-term Cd treatment, Spd application caused no significant change of SOD, POD and CAT activity in Cd-stressed plants, whereas long-term Cd + Spd treatment obviously resulted in the enhancement of both POD and CAT activity compared to those in the Cd treatment group (P < 0.05).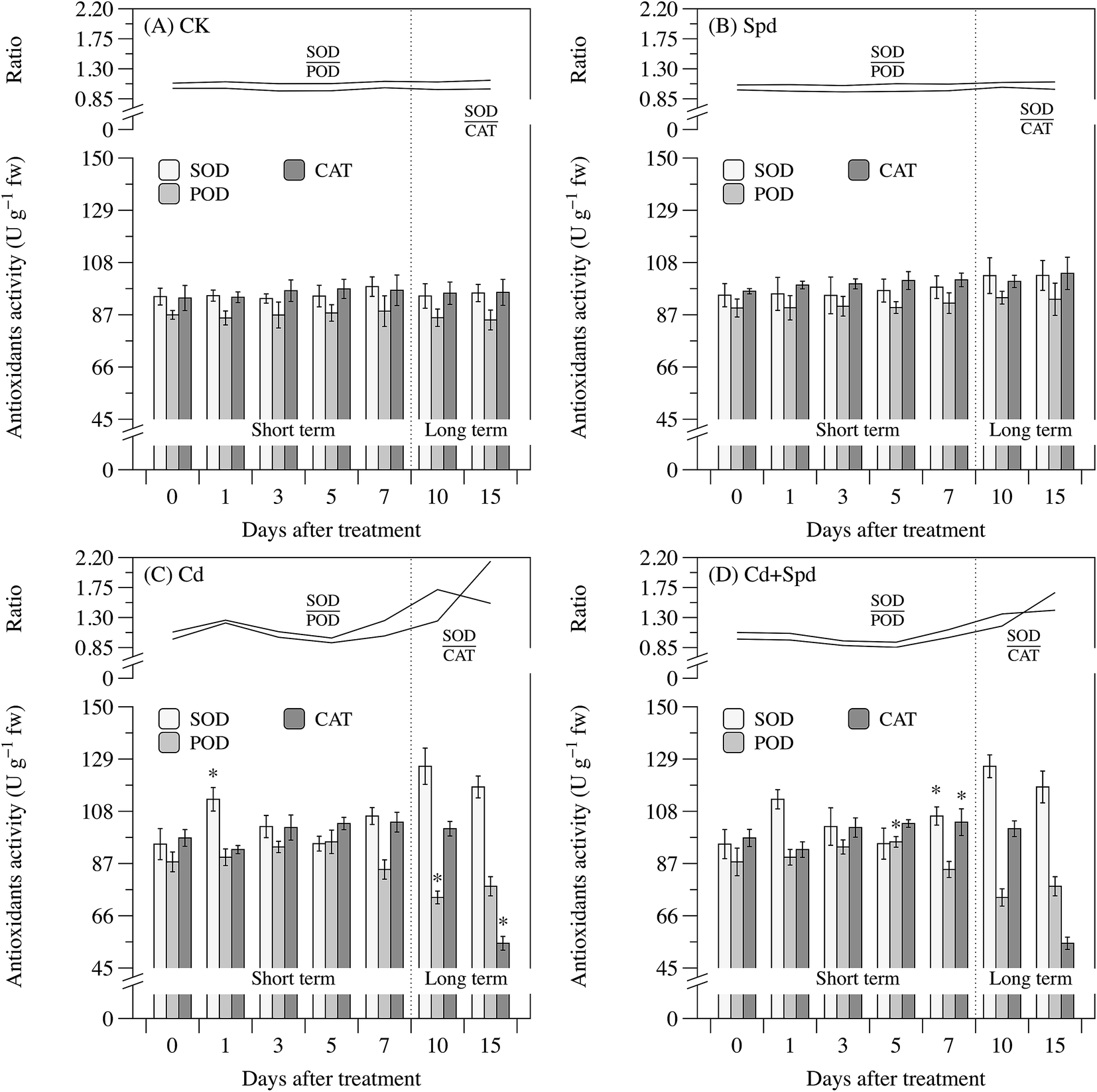 | ||
| Fig. 4 The activities of superoxide (SOD), peroxidase (POD) and catalase (CAT) in the leaves of ramie seedlings after different periods of treatment; (A) 10% Hoagland solution (CK); (B) 10% Hoagland solution + 0.1 mM spermidine (Spd); (C) 10% Hoagland solution + 30 μM cadmium (Cd); (D) 10% Hoagland solution + 30 μM cadmium + 0.1 mM spermidine (Cd + Spd). Each data represents means ± SD of three replicates. Asterisks (*) indicate significant differences (P < 0.05) among the four treatments. | ||
Under Cd treatment, the ratio of SOD activity to POD activity (SOD/POD) and SOD activity to CAT activity (SOD/CAT) slightly increased at early time point, and then maintained a stable trend over the stage from 1 to 5 d. Both of SOD/POD and SOD/CAT were markedly elevated by Cd exposure after 5 d, reaching the highest peak at 10 and 15 d, about 1.6-fold and 2.2-fold of those in the CK plants, respectively. Spd pretreatment caused much lower SOD/POD and SOD/CAT value in ramie leaves compared to Cd stress alone, despite a slight induction of that in non-stressed plants.
MDH is a key enzyme involved in redox status and the lapse of Cd treatment time witnessed its activity fluctuation (Fig. 5A). MDH activity showed a rising trend with the increase in Cd exposure time, and reached the maximum at 7 d, when the activity of MDH was 2.1-fold compared to the CK group. Then, MDH activity collapsed below the CK level after 7 d. The MDH activity in the Cd + Spd treatment group was growing at a much lower rate from 1 to 7 d compared to the Cd treatment group. Spd pretreatment significantly mitigated the effect of long-term Cd stress on MDH activity in leaves (P < 0.05).
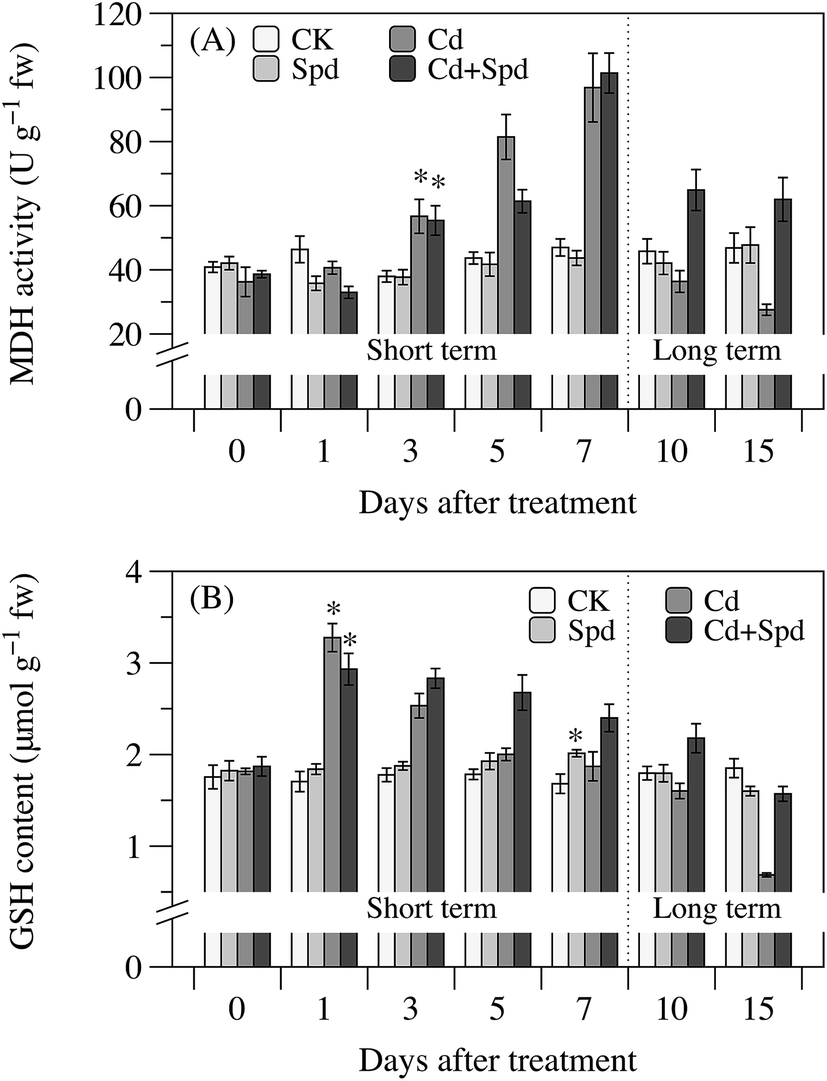 | ||
| Fig. 5 Effects of cadmium (Cd) and spermidine (Spd) application on malate dehydrogenase (MDH) activity (A) and reduced glutathione (GSH) content (B) in the leaves of ramie seedlings after different periods. Each data represents means ± SD of three replicates. Asterisks (*) indicate significant differences (P < 0.05) among the four treatments. CK, Spd, Cd and Cd + Spd correspond to 10% Hoagland solution (HS), HS + 0.1 mM Spd, HS + 30 μM Cd and HS + 30 μM Cd + 0.1 mM Spd, respectively. | ||
3.6. Non-enzymatic metabolite
GSH is the main thiol-containing antioxidant within the cell system. As shown in Fig. 5B, Cd or/and Spd treatments exhibited significant impact on the content of GSH in ramie leaves compared to the control samples (P < 0.05). Higher GSH content was observed in ramies with Spd pretreatment than that in the CK plants. For Cd stress, there was an irregular change in GSH content in the course of time. After Cd disposal, the GSH content increased immediately and reached the peak of 3.3 μmol g−1 at 1 d. However, over the stage from 1 to 15 d, it gradually went downward. The minimum of GSH content, which was 37% of the CK level, was detected at 15 d. Spd pretreatment caused much more GSH synthesis in Cd-stressed ramie leaves at most of the time points tested. After dramatic elevation at 1 d, GSH content in leaves of the Cd + Spd treatment group gradually decreased to the similar level of the CK plants.4. Discussion
The Cd toxicity was affected not only by supplied Cd concentration and duration but also by the amount of available protective compounds induced by Cd exposure. At the optimum condition, Cd could play a positive role in promoting plant metabolism, growth and development, while severe Cd stress may lead to oxidative damage and cellular death. Recent studies have shown that PAs can protect plants from various types of abiotic stress at low concentrations. In the present study, a series of experiments were performed to monitor time-course changes in Cd accumulation, plant growth, photosynthesis, macromolecules metabolism, lipid peroxidation and antioxidative mechanism as well as the influence of Spd pretreatment.The most common symptom of Cd-induced phytotoxicity is growth inhibition, which could be mainly due to the suppression in cell division and elongation.27 In the present study, exposure to long-term Cd stress resulted in the decrease in root and shoot biomass of ramie seedlings, which is an external marker of metabolic disorder inside plants. The results were essentially the same as those in peanut, barley, bean and mustard plants.28–31 Ameliorative impact of exogenous Spd on the growth of ramie seedlings under Cd stress was observed, which may be related to the role of Spd in metal accumulation, ion homeostasis, photosynthesis capacity, membrane integrity and cellular metabolic activity. As shown in Table 1, Cd accumulation in the roots was much higher than that in the above-ground parts (shoots and leaves) after 15 d of Cd exposure. This response suggested a low transport rate of Cd within the ramie, which may be a natural protective measure of plants to defend the aerial parts from Cd toxicity.32 The role of PAs as metal chelating agent has been known, and therefore it could reduce the heavy metal accumulation in each organ and improve the metal transfer efficiency within plants.10 In this study, we found that Spd application decreased Cd uptake and promoted Cd translocation from roots to aerial parts, thus alleviating Cd toxicity to ramie roots and further enhancing the Cd tolerance of ramie.
The hitherto studies showed that Cd may result in two kinds of unfavorable consequences in plant cells, namely (1) the oxidation of macromolecules (chlorophyll, proteins, lipids, carbohydrates etc.) and redox imbalance evoked by the stimulation of ROS formation, and (2) the alteration of component structure or inhibition of enzyme activities by binding of Cd to specific groups (e.g. –SH, –PO4) of proteins and lipids.33 Chlorophyll loss, plant senescence indicator, was observed as a common consequence in plants exposed to heavy metal.34 Accordingly, short-term Cd treatment showed promoting effect on the photosynthetic pigment contents, which could draw a conclusion that ramie seedlings can tolerate Cd at applied concentration without notable leaf necrosis and chlorosis during a certain time span. Then, a decline in the level of chlorophyll and carotenoid were observed after 10 d of Cd exposure, which could be due to the substitution of Mg2+ located in the tetrapyrrole ring of chlorophyll molecules and the inhibition of the enzyme activity related to Calvin cycle or thylakoid membrane peroxidation.33 The ratio of Chl-a to Chl-b may be considered as a marker for the plant's capacity of solar energy utilization. The lower Chl-a/Chl-b ratio in the Cd treatment group at long-term points also indicated oxidative stress and damage to the chloroplast structure.33,35 PAs treatment resulted in the promotion of growth in plants exposed to various abiotic stresses with an associated elevation of pigment contents and photosynthetic efficiency.21 In this study, Spd notably prevented pigment degradation under long-term Cd stress and enhanced the efficiency of solar energy conversion. The proposed mechanism is based on the association of Spd with negatively-charged phospholipids or proteins resulting in the stabilization of molecular structure of photosynthetic apparatus like thylakoid membrane and delay of senescence.21,36 Moreover, carotenoid also serves as an efficient scavenger of singlet oxygen and triplet chlorophyll. Thus elevated amounts of carotenoid in Spd-pretreated leaves would strengthen the capacity to quench ROS and prevent oxidative damage.37 Soluble protein and sugar have been widely studied to be involved in cellular metabolic stability, and accumulations of these essential metabolites are positively correlated with tolerance to oxidative stress. Among them, soluble sugar not only directly quenches ROS by feeding NADPH-producing metabolic pathways, but also induces the gene expression of some antioxidative enzymes. It has been reported that glucose or sucrose could enhance the activity of SOD, ascorbate peroxidase (APX), dehydroascorbate reductase (DHAR) and the ascorbic acid level in plants.38 In this study, Cd exposure caused a statistically non-significant promotion of soluble protein content after 7 d, which is likely due to the induction of stress proteins.39 In contrast, Cd treatment aroused an evident accumulation of soluble sugar at 3 d, therefore contributing to improving antioxidative capacity in advance. However, critical oxidative stress conditions triggered an increase in soluble protein and sugar turnover or degradation, which may result from an oxidative damage of molecular structure and the enhanced proteolytic activity caused by Cd stress.40 The tendency of depletion in soluble protein and sugar, similar to Lupinus albus, was significantly retrieved by Spd application.41 Spd may serve as protein and sugar structure stabilizer, thus allowing ramie seedlings to maximize their nutrient storage reserves and maintain normal intracellular metabolism.42 Hence, short-term Cd stress could promote the basal metabolism of ramie cells; nevertheless, long-term Cd exposure resulted in the chlorophyll loss, soluble sugar and protein degradation. The use of exogenous Spd enhanced the physiological characteristics of Cd-treated ramie seedlings by increasing both pigments and fundamental macromolecules.
MDA is a cytotoxic peroxidation product of polyunsaturated fatty acids of bio-membrane; it is usually regarded as a biomarker of free radical production and consequent oxidative stress.6 In this study, notably, the MDA level showed declining trend when ramie was exposed to Cd stress after 3 d. It suggested that short-term Cd exposure did not arouse serious oxidative stress. In addition, it might also be attributable to the instability of lipid peroxidation in non-enzymatic reactions, the changed mode of lipid peroxidation induced by fatty acids accumulation or the induction of antioxidation mechanism. A similar phenomenon was also observed in leaves of Arabis paniculata and Phaseolus coccineus at moderate Cd level.43,44 With the increased treatment time, the MDA content reached the maximum value at 15 d, which indicated that the prevalence of oxidative stress and membrane deterioration, probably resulted in plasma membrane dysfunction by altering the membrane lipids composition, disrupted the normal ion exchange and the selective uptake of the cells, affected the inside osmotic pressure and finally enhanced Cd accumulation. The increased lipid peroxidation induced by Cd has also been observed in other plant species like rice, mustard and kenaf.4,31,45 The beneficial effect of Spd application was found under long-term Cd exposure. Spd could act as a protector for the plasma membrane by modulating the compartmentalization of Cd and the ions channel of membrane, maintaining membrane integrity through interaction with phospholipids or proteins.46 Under short-term Cd stress, however, the protective effect of Spd was not statistically as significant as that in long-term processing, which supported the view that ramie seedlings could tolerate short-term Cd stress without Spd recovery.
Cell antioxidants play vital roles in plant tolerance to heavy metal stress by protecting labile macromolecules against the attack from free radicals and their variations reflect the ROS formation or the modified redox state in the stressed cells.33 The antioxidative defense system consists of antioxidant enzymes and antioxidant metabolites, whereas, the stress-dependent alterations in these antioxidants were relied on the plant species, Cd concentration used and the time points detected in the present study.47 The first group of antioxidant enzymes involved in the defense system is the ROS-scavenging enzymes including SOD, POD and CAT.48 A significant increase in SOD activity was observed after 1 d of Cd exposure, whereas both POD and CAT activity showed no marked response to the short-term Cd stress. These results are in line with the suggestion of previous studies that SOD constitute the frontline of defense against ROS outbreak.33 The enhanced activity of SOD implied the efficient decomposition of O2˙− into H2O2, hence decreasing the risk of the formation of hydroxyl radicals from O2˙− via Haber–Weiss reaction.49 POD and CAT were responsible for removing H2O2 produced by SOD-ruled dismutation.45 Data presented in this research illustrated that long-term Cd stress triggered down-regulation of POD and CAT activity, whereas no clear decline on SOD activity was found during Cd processing. Thus the ratio of SOD/POD and SOD/CAT which constantly increased over time could indirectly reflect the intracellular H2O2 level or the degree of oxidative stress. The above results can be a confirmation of that long-term Cd stress caused mass H2O2 accumulation and immediate oxidative damage. The second group of enzymes could be activated to generate reducing power by NAD(P)H carriers for various metabolic pathways, thereby regulating the homeostasis equilibrium of redox status in plant cells.48 MDH, a typical NAD(P)+-reducing enzyme, was significantly activated after 3 d of Cd exposure, which is similar to that in Phaseolus vulgaris and pepper plants.48,50 It appeared to be an adaptational trait of Cd tolerant cultivars in maintaining relative stability of redox status and possibly promoting effective operation of TCA cycle.51 Likewise, MDH activity had a critical decline upon long-term Cd stress, which contributed to disrupting the cellular redox state and consequently inhibiting both photosynthesis and respiration. GSH, a precursor of phytochelatins, is the most crucial non-enzymatic antioxidant in plants.40 The GSH level reached the maximum after 1 d of Cd stress, which is correlated with oxidative stress tolerance and redox buffer in ramies. Increase in Cd-stressed time showed a gradual depletion in GSH level, which may be attributed to phytochelatin synthesis, direct neutralization of Cd-induced ROS by creating the oxidised glutathione disulphide (GSSG) or an increased utilization for ascorbate synthesis via ASA–GSH cycle.2 GSH with higher level compared to the controls also played an indirect role in protecting plasma membrane by maintaining a reduced state and preventing protein denaturation induced by the oxidation of thiol groups.48 The GSH content sharply reduced to a lower level than control till the 15th day, which matched the results that long-term Cd exposure leads to membrane peroxidation and protein degradation. It has been shown that Spd application was able to alleviate oxidative damage by altering the antioxidant defense system and ROS level under metal-stressed conditions. During short-term Cd processing, there were no obvious variation in SOD, POD and CAT activity when Spd was added to the medium. By contrast, Spd application decreased the ascending rate of MDH activity and enhanced GSH content in the Cd-stressed leaves. It might suggest that Spd has a redox status maintaining effect by direct scavenging of free radicals, counteracting the Cd-induced activation of NAD(P)H oxidases or increasing GSH synthesis, thereby diminishing superoxide generation and reducing the demand for MDH reaction.52,53 The higher POD and CAT activity were observed under long-term Cd + Spd treatment, so that the spare H2O2 in plant cells could be effectively quenched. Long-term Cd-evoked changes, the depletion of MDH activity and GSH content, were also reduced substantially by exogenous Spd application, thus inhibiting the disorder of redox status and intracellular macromolecules oxidation.
5. Conclusions
This study illustrate that ramie can tolerate moderate Cd stress by triggering self-protective mechanism, whereas, severe Cd stress may beyond the limit of its antioxidative capacity and then lead to oxidative damage. Spd pretreatment can effectively alleviate the Cd-induced oxidative stress, and the dominant mechanism of Spd protection is differential between short-term and long-term Cd exposure. In the early stage of Cd toxicity, Spd may strive to maintain redox balance, while in latter stage it can act as an effective stabilizer of fundamental macromolecules and elevate the level of antioxidant system. It suggests that ramie is a promising remediation plant applied in low-concentration Cd contaminated sites. However, for high intensity phytoremediation, Spd application can play a protective role in the promotion of Cd tolerance during different phase.Acknowledgements
This work was funded by the National Natural Science Foundation of China (Grant No. 41271332) and the Science and Technology Planning Project of Hunan Province, China (Grant No. 2012SK2021).References
- S. le Faucheur, F. Schildknecht, R. Behra and L. Sigg, Aquat. Toxicol., 2006, 80, 355–361 CrossRef CAS PubMed.
- M. Hasanuzzaman, M. A. Hossain and M. Fujita, Biol. Trace Elem. Res., 2012, 149, 248–261 CrossRef CAS PubMed.
- S. Clemens, E. J. Kim, D. Neumann and J. I. Schroeder, EMBO J., 1999, 18, 3325–3333 CrossRef CAS PubMed.
- A. Roychoudhury, S. Basu and D. N. Sengupta, Acta Physiol. Plant., 2011, 34, 835–847 CrossRef.
- R. Mittler, Trends Plant Sci., 2002, 7, 405–410 CrossRef CAS.
- D. Del Buono, G. Ioli, L. Nasini and P. Proietti, J. Agric. Food Chem., 2011, 59, 12109–12115 CrossRef CAS PubMed.
- P. L. Gratão, A. Polle, P. J. Lea and R. A. Azevedo, Funct. Plant Biol., 2005, 32, 481–494 CrossRef.
- S. S. Gill and N. Tuteja, Plant Physiol. Biochem., 2010, 48, 909–930 CrossRef CAS PubMed.
- T. Kusano, T. Berberich, C. Tateda and Y. Takahashi, Planta, 2008, 228, 367–381 CrossRef CAS PubMed.
- M. D. Groppa, M. L. Tomaro and M. P. Benavides, BioMetals, 2007, 20, 185–195 CrossRef CAS PubMed.
- X. P. Wen, Y. Ban, H. Inoue, N. Matsuda and T. Moriguchi, Transgenic Res., 2010, 19, 91–103 CrossRef CAS PubMed.
- A. Bouchereau, A. Aziz, F. Larher and J. Martin-Tanguy, Plant Sci., 1999, 140, 103–125 CrossRef CAS.
- M. D. Groppa and M. P. Benavides, Amino Acids, 2008, 34, 35–45 CrossRef CAS PubMed.
- Q. Zhu, D. Huang, S. Liu, Z. Luo, Z. Rao, X. Cao and X. Ren, Plant, Soil Environ., 2013, 2, 57–61 Search PubMed.
- D. Wang, Y. Liu, X. Tan, H. Liu, G. Zeng, X. Hu, H. Jian and Y. Gu, Environ. Sci. Pollut. Res. Int., 2015, 22, 3489–3497 CrossRef CAS PubMed.
- H. Tang, Y. Liu, X. Gong, G. Zeng, B. Zheng, D. Wang, Z. Sun, L. Zhou and X. Zeng, Environ. Sci. Pollut. Res. Int., 2015, 22, 9999–10008 CrossRef CAS PubMed.
- H. Li, Y. Liu, G. Zeng, L. Zhou, X. Wang, Y. Wang, C. Wang, X. Hu and W. Xu, J. Environ. Sci., 2014, 26, 2508–2516 CrossRef PubMed.
- A. Piotrowska-Niczyporuk, A. Bajguz, E. Zambrzycka and B. Godlewska-Zylkiewicz, Plant Physiol. Biochem., 2012, 52, 52–65 CrossRef CAS PubMed.
- B. Suresh and G. Ravishankar, Crit. Rev. Biotechnol., 2004, 24, 97–124 CrossRef CAS PubMed.
- J. Duan, J. Li, S. Guo and Y. Kang, J. Plant Physiol., 2008, 165, 1620–1635 CrossRef CAS PubMed.
- S. Parvin, O. R. Lee, G. Sathiyaraj, A. Khorolragchaa, Y. J. Kim and D. C. Yang, Gene, 2014, 537, 70–78 CrossRef CAS PubMed.
- L. L. Knudson, T. W. Tibbitts and G. E. Edwards, Plant Physiol., 1977, 60, 606–608 CrossRef CAS PubMed.
- R. L. Heath and L. Packer, Arch. Biochem. Biophys., 1968, 125, 189–198 CrossRef CAS.
- M. M. Bradford, Anal. Biochem., 1976, 72, 248–254 CrossRef CAS.
- E. Yemm and A. Willis, Biochem. J., 1954, 57, 508 CrossRef CAS.
- G. L. Ellman, Arch. Biochem. Biophys., 1959, 82, 70–77 CrossRef CAS.
- D. Liu, W. Jiang and X. Gao, Biol. Plant., 2003, 47, 79–83 CrossRef CAS.
- G. Shi, Q. Cai, C. Liu and L. Wu, Plant Growth Regul., 2010, 61, 45–52 CrossRef CAS.
- A. Metwally, I. Finkemeier, M. Georgi and K.-J. Dietz, Plant Physiol., 2003, 132, 272–281 CrossRef CAS PubMed.
- C. Poschenrieder, B. Gunsé and J. Barceló, Plant Physiol., 1989, 90, 1365–1371 CrossRef CAS PubMed.
- P. Ahmad, G. Nabi and M. Ashraf, S. Afr. J. Bot., 2011, 77, 36–44 CrossRef CAS PubMed.
- S. Singh, S. Eapen and S. F. D'Souza, Chemosphere, 2006, 62, 233–246 CrossRef CAS PubMed.
- D. D. Yilmaz and K. U. Parlak, Ecol. Indic., 2011, 11, 417–423 CrossRef CAS PubMed.
- L. Sandalio, H. Dalurzo, M. Gomez, M. Romero-Puertas and L. Del Rio, J. Exp. Bot., 2001, 52, 2115–2126 CAS.
- D. Horler, J. Barber and A. Barringer, Int. J. Rem. Sens., 1980, 1, 121–136 CrossRef PubMed.
- N. Durmu and A. Kadioğlu, Acta Physiol. Plant., 2005, 27, 515–522 CrossRef.
- B. Demmig-Adams, Biochim. Biophys. Acta, 1990, 1020, 1–24 CrossRef CAS.
- W. Sun, Y. Nie, Y. Gao, A. Dai and J. Bai, Acta Physiol. Plant., 2012, 34, 641–655 CrossRef CAS.
- C. S. Seth, P. K. Chaturvedi and V. Misra, Ecotoxicol. Environ. Saf., 2008, 71, 76–85 CrossRef CAS PubMed.
- R. Szőllősi, I. S. Varga, L. Erdei and E. Mihalik, Ecotoxicol. Environ. Saf., 2009, 72, 1337–1342 CrossRef PubMed.
- G. Costa and E. Spitz, Plant Sci., 1997, 128, 131–140 CrossRef CAS.
- P. Roy, K. Niyogi, D. N. SenGupta and B. Ghosh, Plant Sci., 2005, 168, 583–591 CrossRef CAS PubMed.
- R. Qiu, X. Zhao, Y. Tang, F. Yu and P. Hu, Chemosphere, 2008, 74, 6–12 CrossRef CAS PubMed.
- E. Skórzyńska-Polit and Z. Krupa, Arch. Environ. Contam. Toxicol., 2006, 50, 482–487 CrossRef PubMed.
- F. Li, J. Qi, G. Zhang, L. Lin, P. Fang, A. Tao and J. Xu, Agric. Sci. China, 2013, 12, 610–620 Search PubMed.
- Y. T. Hsu and C. H. Kao, Plant Soil, 2007, 291, 27–37 CrossRef CAS.
- S. S. Sharma and K.-J. Dietz, Trends Plant Sci., 2009, 14, 43–50 CrossRef CAS PubMed.
- K. Smeets, A. Cuypers, A. Lambrechts, B. Semane, P. Hoet, A. van Laere and J. Vangronsveld, Plant Physiol. Biochem., 2005, 43, 437–444 CrossRef CAS PubMed.
- Q. Shi and Z. Zhu, Environ. Exp. Bot., 2008, 63, 317–326 CrossRef CAS PubMed.
- A. M. León, J. M. Palma, F. J. Corpas, M. Gómez, M. C. Romero-Puertas, D. Chatterjee, R. M. Mateos, A. Luis and L. M. Sandalio, Plant Physiol. Biochem., 2002, 40, 813–820 CrossRef.
- R. G. Kumar, K. Shah and R. Dubey, Plant Sci., 2000, 156, 23–34 CrossRef CAS.
- W. Shen, K. Nada and S. Tachibana, Plant Physiol., 2000, 124, 431–440 CrossRef CAS PubMed.
- J. Kubiś, J. Plant Physiol., 2008, 165, 397–406 CrossRef PubMed.
| This journal is © The Royal Society of Chemistry 2015 |