
Effect of freezing methods on sulforaphane formation in broccoli sprouts
Abstract
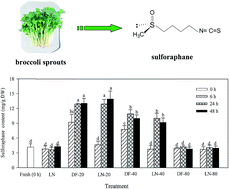
Broccoli sprouts were directly frozen at −20 °C (DF-20), −40 °C (DF-40) and −80 °C (DF-80) or stored at −20 °C (LN-20), −40 °C (LN-40) and −80 °C (LN-80) after being frozen in liquid nitrogen for 5 min or always frozen in liquid nitrogen (LN). The effect of these treatments on glucoraphanin and ascorbic acid content, myrosinase activity, sulforaphane and sulforaphane nitrile formation in broccoli sprouts was investigated. The results showed that glucoraphanin content was not significantly affected by freezing. Treatments of DF-20, DF-40, LN-20 and LN-40 for 24 and 48 h enhanced myrosinase activity and decreased the ascorbic acid content. The enhancement of myrosinase activity after freezing was in parallel with more sulforaphane and less sulforaphane nitrile formation. Besides, freezing at −20 °C was more favorable for sulforaphane formation than −40 °C. Directly freezing at −20 °C increased the sulforaphane yield by 1.54–2.11 fold in the broccoli sprouts of the three cultivars investigated in this study.
 Please wait while we load your content...
Please wait while we load your content...

