
Development of an improved E. coli bacterial strain for green and sustainable concrete technology
Manas Sarkara,
Nurul Alama,
Biswadeep Chaudhuria,
Brajadulal Chattopadhyay*a and
Saroj Mandalb
aDepartment of Physics, Jadavpur University, Kolkata – 700032, India. E-mail: bdc_physics@yahoo.co.in; Tel: +91-9433343917
bDepartment of Civil Engineering, Jadavpur University, Kolkata – 700032, India
First published on 25th March 2015
Abstract
Development of smart bioconcrete materials has recently become an emerging area of research for construction. Here, the silica leaching attribute transferred to an E. coli bacterial strain, has been utilized for higher strength and more durable concrete structures. The silica leaching gene was fished out from the BKH2 bacterium (GenBank accession no.: KP231522), amplified by the PCR technique and cloned into E. coli bacteria via a suitable T-vector to develop a bio-engineered E. coli strain. The transformed bacterial cells when incorporated directly into mortar specimens produced high performance biocomposite materials. Improvements on the compressive strength (>30%), ultrasonic pulse velocity (>5%), and decrease in the water absorption capacity were noted in the bacteria amended mortars. FESEM analysis revealed rod-like crystalline structures within the mortar matrices, and XRD analysis confirmed the development of a new silicate phase (gehlenite). The bioengineered E. coli cells can be directly explored for green and sustainable high performance composites in the near future.
1. Introduction
Increasing the compressive strength, rigidity and longevity of concrete structures with the aid of bioengineering techniques has now gained momentum in the construction field. Due to the heterogeneous composition of the principal components, the properties of the final composites can extensively vary.1 Concrete is typically characterized as a material with a high compressive strength but a low tensile strength. The permeability to liquids, consequential corrosion of reinforcement and susceptibility to chemical hazards are the major limitations of concrete structures. Various chemicals, nanomaterials, steel or other reinforcement materials are widely used in concrete to increase its strength and durability.2 In recent times, different types of microbes and their specific enzymes are used in cementitious materials to increase the compressive and tensile strengths of concrete, though no reports are available on the most commonly used E. coli based bioconcrete materials.3–10 Sometimes, this methodology generates self-healing attributes within concrete which have added advantages for the remediation of cracks and fissures inside the concrete structures.11–13Previous studies have reported that the compressive strength and durability of concrete could be increased using an extracellular bioremediase protein (∼28 kDa) isolated from the hot spring bacterium BKH1.7,11 The bacterium BKH1 (Gene Bank accession no.: FJ177512) is a facultative anaerobe and grows gradually (a 6–7 days old growth culture contains 107 cells ml−1) in a semi-synthetic medium at 65 °C and pH 8.14 The bacterium BKH1 is closely related (99.8% homologous) with Thermoanerobacter thermohydrosulfuricus. The bioremediase protein is a unique thermostable and high-pH tolerant protein secreted by the bacteria in the growth medium and possesses silica leaching activity.14 The amino acid sequence of the protein shows 78% homology with bovine carbonic anhydrase II but there is no functional similarity between them (communicated).
The aim of the present investigation is to explore the most common E. coli bacterial strain having similar characteristics to the hot spring microbial consortium of being industrially viable and commercially applicable in the development of high performance concretes. This study will also facilitate to develop a modified bacterial strain which will be grown quickly at ambient temperature and normal pH (7.0) in a simplified cost effective medium. Here, a genetic manipulation technique has been utilized for developing new characteristics within the E. coli bacteria (JMJ107) by transferring the silica leaching gene from BKH2. The gene was transformed into a host E. coli strain by an appropriate T-vector, and the transformed cells were incorporated into cementitious mortar to observe their usefulness regarding the compressive strength and durability of the amended material. It has already been observed that E. coli itself does not possess the attribute to increase the compressive strength.7 This easy and comparatively low-cost technology for developing high performance concrete materials might be useful for the construction industries where high-quality construction materials are needed for buildings, bridges and the conservation of historical monuments.
2. Materials & methods
2.1. Reagents
All the analytical grade chemicals used in this investigation were purchased from the Sigma Chemical Company of USA, Merck of Germany and Spectrochem Pvt., Ltd. of India. For concrete/mortar sample preparation ordinary Pozzolana Cement 43 grade15 and standard Ennore sand16 were used. A E. coli bacterial strain (JMJ107; MTCC no. 1669) received from ICGEB, New Delhi, India, was used in this study.2.2. Isolation and molecular identification of thermophilic microorganisms for biosilicification
Mixed populations of thermophilic microorganisms were collected from the soil of one of the hot springs (temperature 65 °C) at Bakreshwar, West Bengal, India (23.8800° N, 87.3700° E). An enrichment culture was developed by growing the mixed population culture in a semi synthetic growth medium containing 0.1 M Fe(OH)3, 0.6 g L−1 Na2HPO4, 0.33 g L−1 KCl, 2.5 g L−1 Na2CO3, 0.02% yeast extract and 0.5% peptone at pH 8 and 65 °C under anaerobic conditions for 7 days as described earlier.7 Isolation of the pure bacterial strain from the enrichment culture was done using the serial dilution technique. The photomicrograph of the isolated bacterium was taken by high resolution transmission electron microscopy (HRTEM; JEOL, JEM 2100). Gram staining was performed to identify the nature of the bacterium. The silica leaching attribute of the isolated strain was confirmed by biosilicification assay of the supernatant (taken from the 7 day old cultured medium) against tetraethyl orthosilicate (TEOS) as described previously.14The 16S-rRNA gene sequence primers (forward-5′CAGGCCTAACACATGCAAGTC3′ and reverse-5′GGCGGTGTGTACAAGGC 3′) were used for molecular characterisation. The partial sequence of 16S-RNA was obtained and submitted to the GenBank. For the strain identification the phylogenetic tree of the bacterium was developed, and it was named BKH2.
2.3. Strength measurement
Mortar samples were prepared by incorporating different cell concentrations (101–107 cells per ml water used) of the isolated strain BKH2 into the cement sand mixture to observe the strength-increasing properties. A water vs. cement ratio of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :0.4 and a cement vs. sand ratio of 1:3 were used. The compressive strengths of the bacteria amended samples were measured by a digital compression test machine after different numbers of days (3, 7, 14 and 28) of air curing and water curing (deionized water) as described earlier.17
:0.4 and a cement vs. sand ratio of 1:3 were used. The compressive strengths of the bacteria amended samples were measured by a digital compression test machine after different numbers of days (3, 7, 14 and 28) of air curing and water curing (deionized water) as described earlier.17
2.4. Protein isolation and purification
The bacterium BKH2 secretes few proteins in the growth medium which were purified individually by Sephadex G-100 column chromatography and analyzed for biosilicification activity.14 A protein having silica leaching activity like bioremediase was targeted for the gene of our interest. The protein was purified and further applied in mortar samples to observe the strength-increasing attribute.2.5. Cloning of the gene that codes for the silica leaching protein
Genomic DNA was isolated from the culture pellet of the BKH2 bacterium. Two sets of primers including cloning (restriction) sites for NdeI and XhoI were designed for the study. One set of primer was designed decoding the amino acid sequence with a codon preference for the Thermoanerobacter thermohydrosulfuricus which was closely related to the BKH1 bacterium.14 Primer sequences:Forward primer: AGCTCACATATGTCTCATCATTGGGGATATGG
Reverse primer: AGCACTCTCGAGTTGTGCAGGTCTCCAATTTGCTAAC
The second set was designed with respect to carbonic anhydrase II of Bos Taurus as because, bioremediase protein was found to be 78% homologous to it [communicated].
Primer sequences:
Forward primer: AGCTCACATATGTCCCATCACTGGGGATACGGC
Reverse primer: AGCACTCTCGAGTTTGGGGAAACCTCTGACTTGTC
The following conditions were maintained for the polymerase chain reaction (PCR) in this study:
Initial denaturation: 94 °C – 5′
Denaturation: 94 °C – 30′′
Annealing: 50 °C – 30′′
Extension: 72 °C – 1′
Final extension: 72 °C – 15′
The gene was amplified with both sets of primers. Two clones were obtained from two primers but the clone (∼800 bp) obtained from the second set of primers (FPO/RPO of carbonic anhydrase II of Bos Taurus) and whose product showed biosilicification activity was processed for cloning into the T-vector. Furthermore, the positive clones were screened by digesting with NdeI and XhoI to see a release of 800 bp. The clone DNA was purified and sent for sequencing. The sequence data obtained from the clone was analysed and found to match with the outer membrane porin protein of Cupriavidus metallidurans. The clone was ligated to the T-vector and transformed into the E. coli (JMJ107) strain through a standard transformation process.18
2.6. Expression and identification of cloned gene for biosilicification
The transformed JMJ107 and host cell JMJ107 (E. coli strain) were grown in the Luria Bertani (LB) medium at 37 °C overnight using ampicillin wherever necessary. All cell bacterial proteins were isolated by lysing the cells of both cultures (host and transformed) and run on different lanes of a SDS page electrophoresis gel along with a Sigma protein marker. The desired protein of interest was purified using the Sephadex G-100 gel filtration chromatographic technique. The activity of the purified enzyme was confirmed by biosilicification assay as described by Biswas et al.142.7. Mechanical characterization of bio-mortar samples
Control mortar samples (dimension: 70.6 mm × 70.6 mm × 70.6 mm) were prepared by mixing Ordinary 43 grade Portland Cement, Ennore sand and deionised water as per IS 4031-1988.7 The cement:sand and cement:water ratios were similarly maintained as described earlier in section 2.3 of the materials and methods. The LB medium, host cell JMJ107, transformed JMJ107 and BKH2 were used in the cement–sand–water mixture in the same ratio mentioned earlier for preparing the test mortar specimens. The concentration of BKH2 cells in the test mortar specimens was varied from 101 to 108 cells per ml water used. Whereas three different cell concentrations (107, 108 and 109 cells per ml water used) of host JMJ107 and transformed JMJ107 were used for the test sample preparation. The bacterial cell concentrations were determined directly by noting the optical density (OD) of the bacteria growing cultured media at 620 nm and then by comparing the observed OD with the pre-calibrated OD vs. bacterial cell number calibration plot. Serial dilution for the required bacterial cell concentrations was done by adding the required volume of deionized water to the bacteria-incorporated growth medium. The required volume of water was replaced by LB medium only in the case of LB medium-based sample preparation. The mortar samples were cured under air and water for 3, 7, 14, and 28 days at room temperature to observe the gradual development of various mechanical properties within the bio-mortar samples. A digital compressive strength testing machine (rate 0.5 kN s−1) and Pundit plus PC1007 UPV meter were used to determine the compressive strength and ultra-pulse velocity of those samples as per ASTM C597-02.19,20 Water absorption capacities of the mortar samples were also determined using Neville’s method.21
2.8. Microstructure analysis of bio-mortar samples
After 28 days water curing, the control and transformed bacteria amended mortar samples were broken and individually crushed into fine powder by a pestle-mortar. A little amount of each powder sample was taken, dispersed in absolute ethyl alcohol and dried on a carbon tape in a vacuum desiccator. The dried powder sample was examined by field emission scanning electron microscopy (INSPECT F50 SEM, FEI Europe BV, The Netherlands) and energy dispersive spectroscopy (EDS) coupled with FESEM for structural and elemental analysis.A pinch of the dry powder samples, taken from the JMJ107- and transformed JMJ107 bacteria-incorporated mortar specimens, were sieved to achieve a uniform particle size (∼5 μm) for XRD (Bruker AXS, Inc., Model D8, WI, USA) analysis. The experiment was conducted at 40 kV with a scan speed of 0.5 s per step. The XRD spectra were taken from 2θ = 10 to 80°. The peaks at new positions in the spectrum were marked and analysed using the JCPDS data file.
2.9. Statistical analysis
For each category of testing, five samples were tested. Each experiment was repeated at least three times. Data were presented as average (over 15 samples) with ±SD.3. Results
3.1. Identification of the bacterial strain
A novel anaerobic gram negative and rod-shaped (2–3 μm long and 0.6–0.7 μm width) bacterium (BKH2) was isolated from the hot spring water of Bakreshwar, West Bengal, India, and morphologically identified as small and rod-shaped by HRTEM (Fig. 1). This bacterium was found to grow well at 65 °C (optimum temperature) in a specific enrichment medium. The strain was also processed for 16S based bacterial identification to confirm its authenticity with primers used for 16S-rRNA amplification. The partial 16S-rRNA gene sequence (961 nucleotides) of the bacterium suggested that the isolate belongs to the ‘Burkholderiaceae’ family with 97% identity to the bacterium ‘Cupriavidus metallidurans’ (Fig. 2) as analysed by NCBI BLAST. The phylogenetic tree was developed on the basis of the BLAST results. The bacterium was submitted to NCBI and named BKH2 (GenBank accession no.: KP231522). The silica leaching activity of a protein secreted by BKH2 was confirmed by biosilicification assay (Table 1).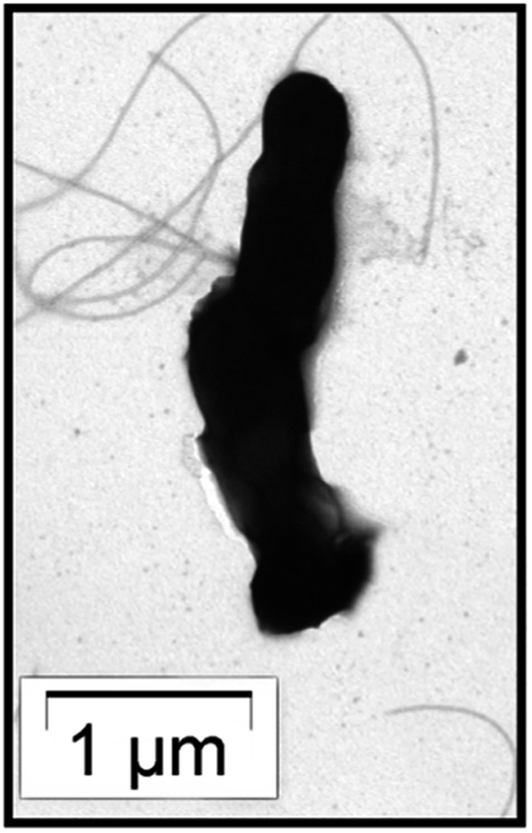 | ||
| Fig. 1 TEM image of the BKH2 bacterium. | ||
 | ||
| Fig. 2 Phylogenetic tree of BKH2 based on the 16S-rRNA (961 nucleotides) sequence. | ||
| Samples | Total protein (mg) | Activity (Units) | Sp. activity (Units per mg) |
|---|---|---|---|
| a One unit of activity of the bioremediase protein is expressed as μg of silica released per mg of protein. | |||
| BKH2 crude protein | 5 | 142 | 28.4 |
| Purified BKH2 protein | 2 | 412 | 206 |
| JMJ107 crude protein | 5 | — | — |
| Transformed JMJ107 crude protein | 5 | 267 | 53.4 |
| Transformed JMJ107 purified protein | 2 | 562 | 281 |
It was observed that the compressive strength of the BKH2 bacteria-incorporated mortar samples increased with the increasing concentration of cells and time as compared to that of the control sample (without bacteria, Fig. 3). The maximum increase in strength was achieved at a bacterial cell concentration of 105 cells per ml water used at all ages (3, 7, 14, and 28 days) when cured under water. More than 30% increase in compressive strength was recorded at 105 bacterial cells per ml of water used under water curing for 28 days.
 | ||
| Fig. 3 Compressive strengths of the BKH2 (101–106 cells per ml water used) amended mortar samples at different days of water curing. | ||
3.2. Isolation and cloning of silica leaching gene
The total genomic DNA of the BKH2 bacterium was isolated and purified. The corresponding gene of the bioremediase-like protein was fished out from the whole genome of BKH2 using the primers constructed from the sequence of carbonic anhydrase II of Bos Taurus (Fig. 4). The DNA fragment obtained using the primers was amplified by the PCR method. The PCR product (clone) was detected on a 1.0% agarose gel containing ethidium bromide and observed by UV transillumination with a Kodak EDAS 290 (Fig. 4). The genomic sequence of the clone (774 bp) was determined by a standard sequencing procedure and is presented in Fig. 5. The isolated clone was then transferred into an E. coli bacterial strain (JMJ107) through a T-vector and the transformation was confirmed by growing the bacterial cells in an ampicillin-containing LB-agar plate, as shown in Fig. 6.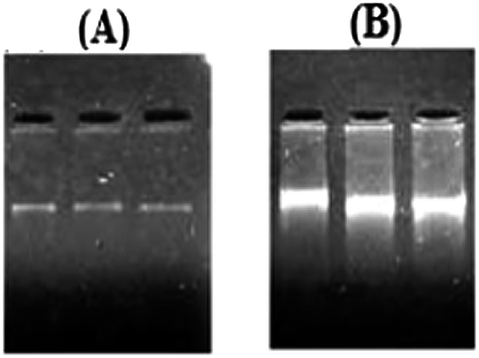 | ||
| Fig. 4 Gel electrophoresis images of the genomic DNA showing (A) the purified genomic DNA of BKH2 and (B) the PCR product of the isolated gene (clone) DNA. | ||
 | ||
| Fig. 5 The sequence of the isolated gene (clone). | ||
 | ||
| Fig. 6 (A) Growth of E. coli (JMJ107 strain) in a LB-agar plate with ampicillin. (B) Growth of transformed E. coli (JMJ107 strain) in a LB-agar plate with ampicillin. | ||
3.3. Expression of the bioremediase-like protein
Expression of the bioremediase-like protein (M.W. ∼28 kDa) in the transformed JMJ107 strain is shown in Fig. 7A. The protein was purified through the Sephadex G-100 column chromatographic technique both from the medium of the BKH2 growth culture as well as from the whole cell lysate of transformed JMJ107 (Fig. 7B).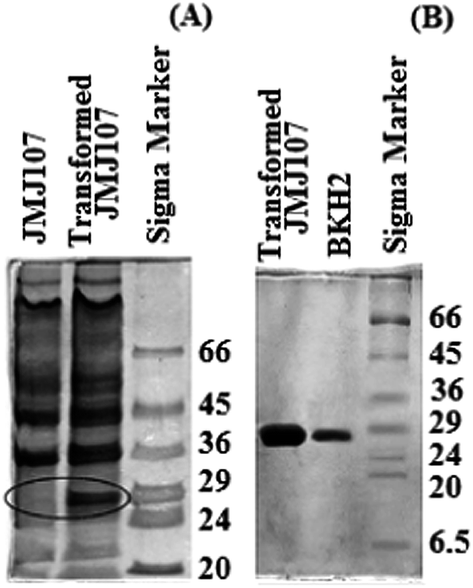 | ||
| Fig. 7 SDS-PAGE images of the protein profiles. (A) Lane 1 – E. coli (JMJ107), Lane 2 – transformed E. coli (JMJ107), Lane 3 – Marker (Sigma). (B) Lane 1 – purified protein of transformed JMJ107, Lane 2 – purified protein of BKH2; Lane 3 – Marker (Sigma). | ||
3.4. Evaluation of mechanical properties of bacteria-incorporated mortar
Transformed JMJ107 cells showed positive strength-increasing properties of the mortar specimens when incorporated into the samples (Fig. 8A and B). The best results for compressive strength were achieved when the transformed cells were incorporated at a concentration of 108 cells per ml of water used (Fig. 8). Fig. 9A and B show the compressive strengths of the mortar samples containing BKH2 and transformed JMJ107 bacterial cells under air and water curing, respectively. The transformed JMJ107 strain achieved a better compactness of the mortar samples compared to those of the control and also with the BKH2 strain, as revealed by the ultra-pulse velocity test (Fig. 10). A 6% increase in ultrasonic pulse velocity was noted for the transformed JMJ107 (T-JMJ107) cell-incorporated mortar samples compared to the control samples. Least water absorption capacity (6.57% increase) was found for the JMJ107 bacteria-modified mortar samples compared to those of the control samples (Table 2). The biosilicification activity of the novel protein expressed by transformation was confirmed and is shown in Table 1.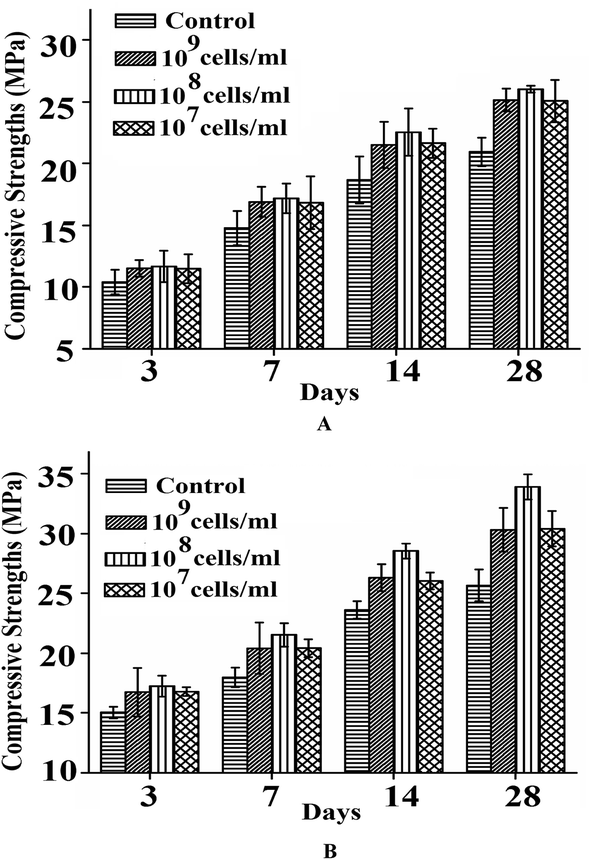 | ||
| Fig. 8 Compressive strengths of transformed JMJ107 amended mortar samples at different days of (a) air curing, and (b) water curing. | ||
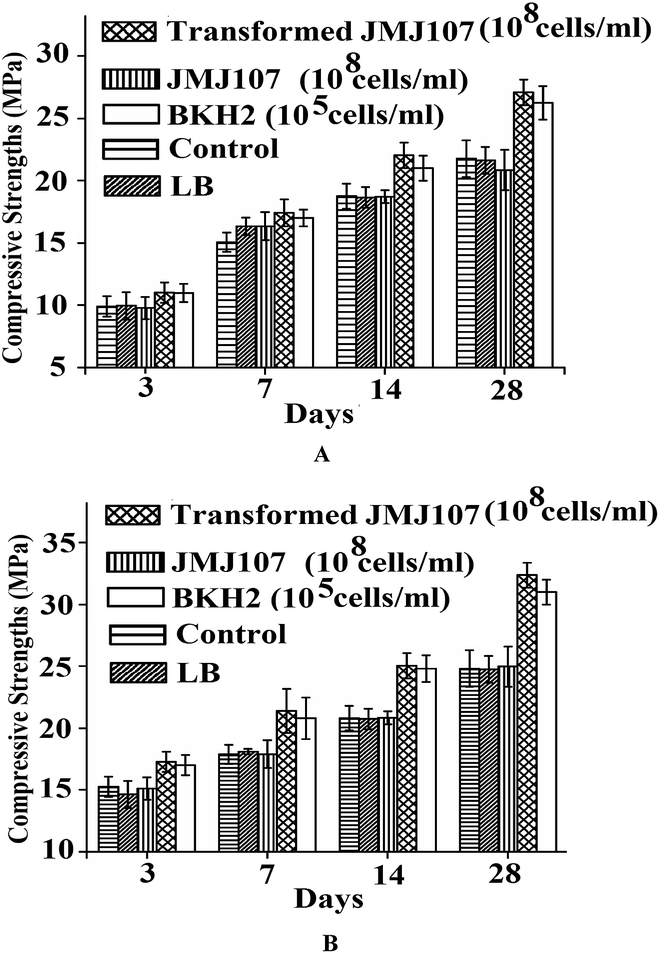 | ||
| Fig. 9 Compressive strengths of the mortar samples containing different bacterial strains under (A) air curing and (B) water curing. | ||
 | ||
| Fig. 10 Ultra-pulse velocity test of mortar samples. | ||
| Sample | Mass (g) | ||||
|---|---|---|---|---|---|
| Initial | After 30 min | % increase | After 24 h | % increase | |
| a Where T-JMJ107 is the transformed bacterium and N = 6. | |||||
| Control | 682.17 ± 0.7 | 700.10 ± 0.2 | 2.63 | 736.17 ± 1.0 | 7.92 |
| LB | 681.83 ± 0.8 | 698.83 ± 0.6 | 2.49 | 735.17 ± 0.6 | 7.82 |
| JMJ107 | 682.93 ± 0.3 | 701.47 ± 0.5 | 2.71 | 736.80 ± 0.8 | 7.88 |
| T-JMJ107 | 664.17 ± 1.9 | 673.14 ± 0.2 | 1.35 | 707.94 ± 1.0 | 6.57 |
| BKH2 | 662.74 ± 0.4 | 672.00 ± 0.3 | 1.39 | 708.94 ± 1.2 | 6.97 |
The microstructures of the different mortar matrices were analysed by FESEM which confirmed the formation of novel irregular nanorod-shaped materials within the transformed JMJ107- and BKH2-containing mortar matrices (Fig. 11). A new phase appeared within the mortar matrices and was confirmed as a calcium–aluminium silicate phase (Gelhenite) by EDS (Fig. 11) and XRD (Fig. 12) analyses.
 | ||
| Fig. 11 Microstructure SEM images of the mortar samples: (A) control sample, (B) LB-treated mortar sample, (C) JMJ107-treated mortar sample, (D) transformed JMJ107-treated sample, (E) BKH2-treated sample. (F) Elemental analysis of the transformed JMJ107 amended mortar matrix by EDS. | ||
 | ||
| Fig. 12 XRD analysis of transformed JMJ107 bacteria amended mortar with respect to host JMJ107 bacteria amended mortar samples. | ||
4. Discussion
The isolated hot spring bacterium (named as BKH2) is a small, rod-shaped and gram negative microbe. The phylogenetic analysis on the basis of 16S-rRNA gene sequences of the newly isolated gene suggests that the bacterium is novel and anaerobic by nature. The bacterial strain is closely related to Cupriavidus metallidurans sp. under the phylum Proteobacteria. The optimum growth conditions for BKH2 have been observed to be 65 °C at pH 8.0 in a specific semi-synthetic medium.14 The bacterium BKH2 is found to increase the compressive strength and durability of the mortar samples when incorporated with different cell concentrations into the samples. It secretes few proteins in the medium during its growth. One of the proteins possesses biosilicification activity, like the bioremediase, which has been confirmed to leach silica from silica-rich substrates (Table 1). The silica leaching behaviour of the protein is responsible for the higher compressive strength as well as more durability as reported earlier.11,14E. coli is the most common and widely used bacterium. It can be cultured easily with short duration and at ambient temperatures in a cost effective manner. This bacterium does not have any strength-increasing properties like BKH1 or BKH2 that can be used for the development of high performance bioconcrete materials. Though the bacteria BKH1 and BKH2 show positive strength-increasing attributes for concrete/mortar samples,7,22 yet the maintenance and growth of these bacteria is a tedious job due to their anaerobic and thermophilic nature. Also, large scale production of these bacteria or their protein need sufficient time as they require 6–7 days of incubation to reach a cell concentration of 107 ml−1 of culture. On the other hand, one can get sufficient E. coli cells (107 ml−1 of culture) within 4–6 h by growing the bacterium in a suitable growth medium (LB medium). Keeping all this in mind, the genetic transformation process has been explored to develop the silica leaching characteristic within the E. coli bacterial strain JMJ107, so that the transformed bacterium can be used in the preparation of bioconcrete materials. The responsible gene that encodes the bioremediase-like protein has been identified from the bacterial BKH2 genome. The gene is then transferred into the E. coli bacterial strain JMJ107 via a suitable T-vector. The antibiotic (ampicillin) resistant nature of the T-vector has been used to ensure the required transformation event. The host E. coli strain fails to grow on an ampicillin-containing LB-agar plate whereas large numbers of distinct colony forming units (CFUs) are observed for the transformed JMJ107 strain under similar conditions (Fig. 6). Positive expression (in terms of protein) of the inserted gene has been observed for the transformed cells (Fig. 7B). The expressed protein similarly shows biosilicification activity as observed from Table 1. The expressed protein (2 μg g−1 cement) similarly shows compressive strength-increasing properties for the mortar samples when incorporated into those (data not presented). Hence, it is possible to get sufficient amounts of protein in shorter time to be used for large amounts of cementitious materials in construction technology.
Compressive strength analysis of the transformed JMJ107-containing mortar samples clearly shows that the transformed bacterial cells can directly be used to significantly increase the compressive strength of the materials (Fig. 8 and 9). A maximum strength increase (>30%) is noted at a cell concentration of 108 cells per ml water used, above which the strength is slightly lowered which may be due to the disruption of the matrix integrity in the presence of excess bacterial cells or protein as explained by Ghsoh et al.22 Interestingly, the transformed JMJ107 strain exhibits better performance regarding the compressive strength than that of the BKH2 strain, as revealed by the results in Fig. 9A and B. In fact, E. coli cannot survive inside the concrete/mortar matrix because of the high alkalinity and anaerobic conditions inside the matrix. In a highly alkaline environment, the bacterial cell wall is disrupted and the protein that is expressed by the inserted gene within the cell against the stress is set free inside the matrices. This protein possesses silica leaching activity and remains active in an environment with very high pH similar to bioremediase.14 The protein is able to produce a new silicate phase that fills the micropores inside the matrices resulting in the enhancement of the compressive strength. Microstructure analysis of the mortar matrices clearly shows the development of novel structures in the transformed JMJ107 and BKH2 bacteria amended samples. The formation of gehlenite (calcium–aluminium silicate) has been confirmed by EDS and XRD analyses. This finding corroborates the result of Sarkar et al.,11 where they have explained the strength-increasing phenomenon of BKH1-containing mortar specimens. The least amount of water absorption (Table 2) and greater ultra-pulse velocity (Fig. 10) also demonstrated that the porosity of the transformed JMJ107-containing cementitious matrices is reduced resulting in an increase in durability of the material.
5. Conclusion
Using microbes has been suggested to enhance the strength of concrete and lower the cement content for making a concrete structure of similar strength. It is established that the microbiologically induced new phase formation or growth inside cement-sand mortar/concrete increases its strength and durability. This means in practice that a substantial part of the cement of mortar/concrete mixtures can be left out while still obtaining the necessary final strength. This would substantially improve the ecological footprint as cement causes massive CO2 emission during its production which affects the global climate negatively. Exploration of lucrative and easy methodologies for the development of bioconcrete materials using the most common E. coli bacterial strain is therefore highly important for commercial purposes which creates new hope for construction technologies.Acknowledgements
Financial assistance for this investigation received from the Department of Biotechnology, New Delhi, Government of India, and their R&D Grant and file number BT/PR-11289/BCE/08/707/2008, dated 30 May 2011, is gratefully acknowledged. The PI is also grateful to DBT for continuing the work in two successive projects.References
- H. M. Jonkers, Self-healing Concrete: A Biological Approach, in Self-healing materials: an alternative approach to 20 centuries of materials science, ed. S. Van der Zwaag, Springer, The Netherlands, 2007, pp. 195–204 Search PubMed.
- P. K. Mehta and P. J. M. Monteiro, in Concrete: Microstructure, Properties, and Materials, ed. P. Hall, New Jersey, 1993 Search PubMed.
- W. C. H. Skinner and H. A. Jahren, in Treatise on geochemistry, ed. W. H. Schlesinger, Elsevier, Holland, 2007, vol. 8.04, pp. 1–69 Search PubMed.
- L. N. Belkova, K. Tazaki, J. R. Zakharova and V. V. Perfenova, Microbiol. Res., 2007, 162(2), 99–107 CrossRef PubMed.
- A. L. Boskey, Biomineralization: an overview, Connect. Tissue Res., 2003, 44, 5–9 CrossRef CAS.
- D. Adak, M. Sarkar and S. Mandal, Construct. Build. Mater., 2014, 70, 453–459 CrossRef PubMed.
- P. Ghosh, S. Mandal, B. D. Chattopadhyay and S. Pal, Cem. Concr. Res., 2005, 35, 1980–1983 CrossRef CAS PubMed.
- S. K. Ramchandran, V. Ramakrishan and S. S. Bang, ACI Mater. J., 2001, 98, 3–9 Search PubMed.
- B. DeGraef, W. DeWindt, J. Dick, W. Verstraete and N. DeBelie, Mater. Struct., 2005, 38, 875–882 CAS.
- S. S. Bang, J. K. Galinat and V. Ramakrishnan, Enzyme Microb. Technol., 2001, 28, 404–409 CrossRef CAS.
- M. Sarkar, T. Chowdhury, B. D. Chattopadhyay, R. Gachhui and S. Mandal, J. Mat. Sci., 2014, 49, 4461–4468 CrossRef CAS.
- H. M. Jonkers, A. Thijssen, G. Muyzer, O. Copuroglu and E. Schlangen, Ecol. Eng., 2010, 36, 230–235 CrossRef PubMed.
- V. Wiktor and H. M. Jonkers, Cem. Concr. Compos., 2011, 33, 763–770 CrossRef CAS PubMed.
- M. Biswas, S. Majumdar, T. Chowdhury, B. D. Chattopadhyay and S. Mandal, Enzyme Microb. Technol., 2010, 46, 581–587 CrossRef CAS PubMed.
- IS 8112: Specification for 43 grade ordinary Portland cement, Bureau of Indian Standard, New Delhi, India, 1989.
- IS 650: Specification for standard sand from natural sources of Ennore, Tamil Nadu District, India for concrete, Bureau of Indian Standard, New Delhi, India, 1991.
- S. Majumdar, M. Sarkar, T. Chowdhury, B. D. Chattopadhyay and S. Mandal, Open J. Civ. Eng., 2012, 2, 218–228 CrossRef.
- D. Hanahan, J. Mol. Biol., 1983, 166, 557–580 CrossRef CAS.
- IS 4031: determination of compressive strength of hydraulic cement, Bureau of Indian Standard, New Delhi, India, 1988.
- ASTM 5977–02: Standard test method for pulse velocity through concrete, ASTM International, West Conshohocken, 2002.
- A. M. Neville, in Properties of Concrete, Pearson Higher Education, Prentice Hall, New Jersey, 1996 Search PubMed.
- S. Ghosh, M. Biswas, B. D. Chattopadhyay and S. Mandal, Cem. Concr. Compos., 2009, 31, 93–98 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2015 |