
Investigation of antiglycation activity of isoprenaline†
Abstract
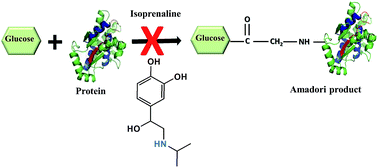
Advanced glycation end products (AGEs) are implicated in pathogenesis of diabetes and its complications. In this study, we report the ability of isoprenaline to inhibit the AGE modification of protein by fluorescence spectroscopy and western blotting. Isoprenaline was more effective in inhibiting AGE modification than aminoguanidine, a well known glycation inhibitor. Further, we show that isoprenaline inhibits at Amadori product formation during glycation reaction by various techniques such as MALDI-TOF-MS, LC-MS/MS, and fructosamine assay. Mass spectrometric analysis of the glycation reaction mixture incubated with isoprenaline suggested that it forms adducts with glucose and thus inhibits glycation. The finding of the additional property of isoprenaline of inhibiting glycation suggests that it is a potential candidate for drug repositioning for the treatment of diabetes and its complications, as it is an FDA approved drug.
 Please wait while we load your content...
Please wait while we load your content...

