
The ORAC (oxygen radical absorbance capacity) index does not reflect the capacity of antioxidants to trap peroxyl radicals
Abstract
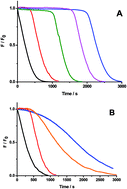
In the present work we demonstrate from kinetic studies that under the experimental conditions proposed for the ORAC protocol, ORAC values do not correlate with the capacity of antioxidants to trap peroxyl radicals (ROO˙), suggesting a dominant role of alkoxyl radicals (RO˙) in the assay.
 Please wait while we load your content...
Please wait while we load your content...

