
Control of Microcystis aeruginosa growth and associated microcystin cyanotoxin remediation by electron beam irradiation (EBI)
Abstract
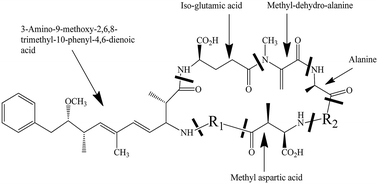
Microcystin-LR (MC-LR), a problematic potent cyanotoxin, is produced by a variety of cyanobacteria. The presence of MC-LR in drinking water is a severe threat to human health as well as an environmental concern. The control of these algal blooms and their associated toxins is critical for ensuring safe drinking water to significant populations. To the best of our knowledge, this is the first detailed study about the application of Electron Beam Irradiation (EBI) for the control of Microcystis aeruginosa algae cultures and simultaneous degradation of MC-LR. The effects of EBI dose on MC production and removal efficiency were investigated by measuring intercellular and extracellular MC concentrations. The dramatic decreases in MC concentration in the cells and solution were observed under our experimental conditions. The correlation between Chl-a and MC concentrations is eliminated. The inhibition of cell growth and degradation of MC-LR by EBI is highly efficient during radiolysis.
 Please wait while we load your content...
Please wait while we load your content...

