DOI:
10.1039/C4RA16649C
(Paper)
RSC Adv., 2015,
5, 29684-29692
Superantigenicity analysis of staphylococcal enterotoxins SElK and SElQ in a mouse model
Received
3rd January 2015
, Accepted 9th March 2015
First published on 16th March 2015
Abstract
Staphylococcal enterotoxins (SEs) are superantigenic toxins secreted by Staphylococcus aureus that is involved in causing food poisoning and human diseases. So far, more than 20 genotypes of SE and SE-like proteins (SEls) have been identified. While many SEs have been found to be capable of causing food poisoning, the role of SEIs in food and human health is still largely unknown. In this study, we analyzed the superantigenic activity of two new types of recombinant SEIs, SElK and SElQ, in a mouse model. The results show that the rSEIK and rSEIQ stimulated distinct murine T-lymphocyte proliferation, caused tumefaction in mouse spleen and thymus, SEls induced increase of cytokines (IL-2, IL-4, IL-6, TNF-α, IFN-γ) measured by quantitative PCR and ELISA both in vitro and in vivo. The results showed that the rSEIQ displayed stronger superantigenicity than the rSEIK and all caused cytokine storm and inflammatory syndromes. The molecular basis for the difference in the superantigenicity was further analyzed by 3D structural modeling. The structural difference (e.g. the critical amino acids of the α3–β8 loop) might partially explain the distinct immune-stimulatory activity of rSElK and rSElQ.
Introduction
Food safety issues caused by bacterial enterotoxin food poisoning have increasingly given rise to public concern. Staphylococcal enterotoxins (SEs), soluble extracellular proteins excreted by Staphylococcus bacteria, have been in a leading role in plenty of grave food poisoning cases. Among all the Staphylococcus bacteria, Staphylococcus aureus is the major one producing a large variety of enterotoxins, which are responsible for infection and intoxication in humans, such as acute gastro-enteritis, Kawasaki-like disease, dermatosis, respiratory diseases, presenting specific acute clinical syndromes like food poisoning.1–4 These enterotoxins share certain genetic characteristics, similar structural and biological functions and exhibit superantigenic activity.5–8 The staphylococcal superantigens act as activators to simulate polyclonal T-cell proliferation through crosslinking T cell receptors (TCRs) with major histocompatibility class II (MHCII) molecules on antigen-presenting cells (APCs). The immune response towards SEs was performed by recognizing specific subtypes of Vβ-TCRs with the outer region of MHCII to activate massive TCRs and APCs and simultaneously cause cytokine storm and inflammatory syndromes.7–12
Up to date, more than 20 genotypes of SE and SE-like genes have been discovered. Some classical types of staphylococcal enterotoxin (SEs, SEA to SEE, SEG to SEI, SER to SET)13,14 are referred to have the capacity to cause food poisoning.15–18 In contrast, although staphylococcal enterotoxin-like proteins (SEl), e.g., SElJ, SElK, SElL, SElM, SElN, SElO, SElP, SElQ, SElS, SElU, SElV and SElT, are homologous and structurally similar to the SEs,13,19,20 little is known about the significance of these SEIs in strains of staphylococcus bacteria from animal infection, and much of the mechanism of the ability of SEs to induce food poisoning remains unknown. Further, their possible role in the development or induction of autoimmune disease has not been described. It has been known that the SE/SEl genes are carried on movable genetic elements (MGE) such as phages (SEA, SEE, SElP), plasmids (SED, SElJ, SElR), and pathogenicity islands (SaPIs),3 which are horizontally transferable among staphylococcal strains. This features bring potential crisis to food industry and human health. Thus SEs have risen a wide range of concerns in human health.
Staphylococcal enterotoxins (SEs), as a kind of virulence factors secreted by S. aureus, are the major decisive causation of staphylococcal food poisoning and the toxic shock symptoms (TSS).21,22 They are well-functioned superantigens defined by their unique ability to systemically alter immune system by affecting T lymphocyte and APCs cytokine production.23 Orwin et al. have examined the biological activities of SElK in the superantigenicity, pyrogenicity, the ability to enhance the lethality in a rabbit model.19,24 However, little is known about the superantigenicity difference among noval SEs and their roles in food poisoning. Thus, we here aim to explore the superantigenic effect of two new types enterotoxins (SElK and SElQ) in a mouse model. The impact on lymphocyte proliferation and cytokine transcription of them were tested in vivo and in vitro. Furthermore, the internal molecular relation within superantigenicity and their structures was elucidated, too.
Materials and methods
Animals
6 week-old C57BL/6J and BALB/C mice were purchased and maintained in SPF facilities in the Institute of Experimental Animal Center, National Academy of Medical Science (Tianjin, China). Animal experiments were performed in compliance with the regulations of Tianjin University Institutional Animal Care and Use Committee (TJIACUC).
Expression and purification of recombinant SElK, SElQ
SElK (accession no.: ABD22279.1) and SElQ (accession no.: ABD21542.1) genes were amplified with primers:
SElK1: 5′-CCGGATCCCAAGGTGATATAGGAATTGA-3′, SElK2: 5′-AACTCGAGTTATATCGTTTCTTTATAAGA-3 (669bp);
SElQ1: 5′-CAGGATCCGATGTAGGGGTAATCAACCTT-3′, SElQ2: 5′-AACTCGAGTTATTCAGTTTTCTCATATGA-3′ (660bp). And the two genes were subsequently cloned to pET28α expression vector.25,26 The positive SEs plasmids (pET28α-SEK, pET28α-SEQ) were constructed and transformed into E. coli BL21(DE3) host cell, and then were induced by 0.1 mM IPTG at 28 °C for 6 hours.27 The recombinant SEs (rSElK, rSElQ) were purified with Ni-NTA purification system (Invitrogen™, USA) according to the manufacture's instruction. SAgs were further purified to homogeneity by thin-layer isoelectric focusing as described as Bao.51 The purified proteins were desalinated and freeze-dried for long-term preservation. Protein quality and bioactivity were analyzed by SDS-PAGE and western bolt according to the previous protocol.27,28
Mouse lymphocyte proliferation by MTT assay
Staphylococcal enterotoxins, rSElK, rSElQ stimulating T cells proliferation29,30 were implemented as follows. Lymphocytes were isolated from thymus of BALB/C or C57BL/6J mice, and seeded in 96-well cell plates at 2 × 105 cells per well in 10% FCS-RPMI-1640 (Gibco, USA). The superantigenic effect was explored by treating lymphocytes with various doses (10, 20, 40, 80 ng mL−1) of rSElK, rSElQ and natural SEA, ConA (10 ng mL−1) as positive control, and PBS buffer as blank control. Four replicates were set for each group with at least three times repeating. Additionally, SEs superantigenic ability was investigated in the presence or without of the A549 cells, a human alveolar adenocarcinoma cell line expressing MHCII molecules on cell surface.31,32 A549 cells were seeded at 1 × 105 cells per well with SEs stimulation and the details were summarized in Table 1. Cell incubation was proceeded at 37 °C with 5% CO2, 95% humidity for 48 h following MTT assay as reported previously.26 Absorbance at 570 nm was measured by a microplate reader. The stimulation index (SI) was calculated by formula: SI = OD570experiment group/OD570blank control and statistically analyzed by Student's t-test.
Table 1 Summary of superantigenic analysis on rSElK, rSElQ treated T cells proliferation compared with inartificial SEA in vitro
| Group |
|
Compositions (total volume 150 μL) |
| Zero adjustment |
|
Culture solution |
| Tumour cell control |
|
Culture solution, A549 |
| Lymphocytes background discharge |
Blank |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes |
| Positive control |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes, 10 ng mL−1 ConA |
| Test of SEs |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes, rSElK, rSElQ, SEA at concentration of 40 ng mL−1, A549 |
| Lymphocytes growth promotion with A549 |
Blank |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes, A549 |
| Positive control |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes, 10 ng mL−1 ConA, A549 |
| Test of SEs |
Culture solution, BALB/C or C57BL/6J thymus lymphocytes, rSElK, rSElQ, SEA at concentration of 40 ng mL−1, A549 |
Mouse viscera index (VI) assay
Six week-old BALB/C mice were randomly divided into four groups (SEA, rSElK, rSElQ and PBS) with 10 mice each. The designated SEs were injected to the mice at a dose of 5 μg kg−1 through caudal vein and 10 mice in each group got same treat for repeat. After 72 h post treatment, the mice were weighed and sacrificed to obtain thymus and spleen. These viscera were weighed and immediately frozen in liquid nitrogen. Mouse viscera index (VI) was figured out by the formula: VI = viscera weight/body weight × 100%.33
Relative Real-Time PCR assay
Total RNAs were extracted from SEs stimulated thymus tissue in vivo and cultured cells in vitro by TRIZOL LS (Promega, USA) according to the manufacture's instruction, and reverse-transcripted by TransScript First-strand cDNA Synthesis kit (TransGen, China). The mRNA transcriptional level of different cytokines (IL-2, IL-4, IL-6, TNF-α and IFN-γ) were detected by relative Real-Time PCR performed on an Applied Biosystems 7500 Real-Time PCR thermocycler. Then the resulting cDNA template were amplified in a 20 μL PCR reaction system34 consist of 100 ng cDNA, 10 μL 2 × SYBR Green mix (Toyobo, Japan), 20 pmol each primer and DNAase-free water. The Real-Time PCR process contained a denaturing step at 95 °C for 3 min, followed by 40 cycles including denaturing at 95 °C for 30 s, annealing at 49.8 °C to 55.9 °C (Table 2) for 60 s and then extension for 30 s at 70 °C, following with melting curve and amplification curve analysis.35 β-Actin used as the endogenous gene and mock treated samples as the calibrator were applied for delta cycle thresholds. The Real-Time PCR data were plotted as the ΔRn fluorescence signal versus the cycle number.36 Cytokines transcription in target cells were measured by relative quantity against β-actin endogenous control by this formula:
where ΔΔCt = (CtTargetx − CtActin)Sampley − (CtTargetx − CtActin)Control.
Table 2 Primers of cytokines for Real-Time PCR
| Gene |
GenBank accession no. |
Primer (5′–3′) |
Annealing Tm (°C) |
Product length (bp) |
| IL-2 |
NM_008366.3 |
(Up)GCGGCATGTTCTGGATTTGACT |
52.5 |
136 |
| (Down)CTCATCATCGAATTGGCACTCA |
| IL-4 |
M25892.1 |
(Up)TCACAGCAACGAAGAACACCAC |
54.6 |
155 |
| (Down)GCATCGAAAAGCCCGAAAGAGT |
| IL-6 |
NM_031168.1 |
(Up)ATGGCAATTCTGATTGTATG |
49.8 |
212 |
| (Down)GACTCTGGCTTTGTCTTTCT |
| IFN-γ |
NM_008337.3 |
(Up)AACTCAAGTGGCATAGATGTGGAAG |
54.1 |
256 |
| (Down)TGTTGACCTCAAACTTGGCAATAC |
| TNF-α |
BC117057.1 |
(Up)TGAGGTCAATCTGCCCAAGTA |
55.7 |
268 |
| (Down)AGGTCACTGTCCCAGCATCT |
| β-actin |
NM_007393.3 |
(Up)AGAGGGAAATCGTGCGTGAC |
55.9 |
204 |
| (Down)CACAGGATTCCATACCCAAG |
ELISA assay
Five cytokines IL-2, IL-4, IL-6, TNF-α and IFN-γ in mice blood and supernatant from cell culture were determined by ELISA kit to reveal expression levels. According to the instructions of mouse cytokine detection kit (R&D, USA), standard curves were constructed as listed below. Simultaneously, the cytokine levels of SEs-treated cell supernatants were measured by this assay.
| y = 0.0029x + 0.044, R2 = 0.9882 (IL-2) |
| y = 0.0059x + 0.035, R2 = 0.9863 (IL-4) |
| y = 0.0077x + 0.0395, R2 = 0.9839 (IL-6) |
| y = 0.0014x + 0.0903, R2 = 0.9879 (TNF-α) |
| y = 0.0006x + 0.089, R2 = 0.9866 (IFN-γ) |
Homology remodeling analysis
The structure was solved by molecular replacement methods using the structure in PDB as a Model. Based on the homologous templates in PDB database, tertiary structure of SElK (PDB ID: 3EA6) and SElQ (PDB ID: 2G9H) have been simulated by homology modeling function of Swiss Model (http://swissmodel.expasy.org/). Signal peptide of staphylococcal enterotoxin proteins was analyzed by online analysis software (http://www.cbs.dtu.dk/services/SignalP/). DNAMAN and spdbv 4.0 software were separately applied to analyze amino acid sequences and spatial structures of the two SEs.
Statistics
The statistical significance of the results were determined using the software GraphPad Prism (version 5.0a; San Diego, CA), and are expressed as the mean ± standard error of the mean. Statistical significance was determined by a Student's t-test, two-way ANOVA for multiple comparisons. Probability values less than 0.05 (p < 0.05) were considered statistically significant.
Results
Expression and purification of recombinant SElK and SElQ
To explore superantigenicity of rSElk and rSElQ, the corresponding genes were individually cloned into bacterial expression vector PET28α with a his-tag gene sequence linked to 5′ end of the enterotoxin gene to promote protein purification. On the basis of superantigenicity potential, natural SEA was chosen as positive control, whose superantigenic ability has been proved previously.37–39 The two target proteins were expressed in E. coli BL21(DE3) cells and purified by nickel column mentioned in previous method. According to SDS-PAGE electrophoresis, the two proteins revealed uniformity in a single band with the expected molecular weights of 26.2, 25.8 kDa (Fig. 1A). Reactogenicity of the two SEls was further verified by western-blot (Fig. 1B).
 |
| | Fig. 1 (A) SDS-PAGE and western-blot analysis of rSElK, rSElQ, and natural SEA, SDS-PAGE electrophoresis of purified proteins rSElK, rSElQ. (B) Western-blot analysis of two rSEs. M stands for protein marker. | |
rSElK and rSElQ differ in ability to cause mouse T cell proliferation
To study the ability of recombinant SElK or SElQ in simulating T cell proliferation, we tested T cells isolated from BALB/C or C57BL/6J mouse thymus. Under different doses of rSElK or rSElQ (10 ng mL−1, 20 ng mL−1, 40 ng mL−1, and 80 ng mL−1) treatment with SEA and PBS as positive and negative controls separately, these cells were evaluated by MTT assay after 48 h post treatment. The stimulation index of 3.627 was gotten from ConA positive treatment group. As shown in Fig. 2, natural SEA, rSElK and rSElQ exhibited a extremely significant dose-dependent stimulation activity compared to PBS control (p < 0.001) and a significant difference (p < 0.05) in different SE treatment groups at the same SE stimulation concentration. SEs can provoke the mechanism of apoptosis,40 high dose (80 ng mL−1) SEs may cause T cell apoptosis. The dose of 40 ng mL−1 rSElK caused significantly more proliferation of lymphocytes than 10 ng mL−1 rSElK in BALB/C mice (p < 0.001).
 |
| | Fig. 2 Different doses of SEls induce distinct T cells proliferation in BALB/C in vitro. MTT assay was carried out in 96 plates at concentrations of 10 ng mL−1, 20 ng mL−1, 40 ng mL−1, 80 ng mL−1 covering two rSEls (rSElK, rSElQ), natural SEA (positive control) and negative control PBS. Y axis was the stimulation index, which indicates the value in treatment groups compared to PBS control. SI > 1 indicates an increase in lymphocyte proliferation after treatment. Error bars represent the standard deviations, and statistical significance was determined by using Student's unpaired t-test. | |
TCR expressed on T cells crosslinking with MHC II on antigen presenting cells by superantigen enhance the T cell proliferation. To verify whether co-incubation of T cell with APC cells boost up the numbers of T cell proliferation, we incubated BALB/C or C57BL/6JT mouse thymocytes with a cell line expressing MHC II molecules, A549 cells and stimulated the mixture with 40 ng mL−1 rSEls. MTT assay was implemented after 48 h post treatment. Compared with negative control, the presence of A549 cells significantly increased (p < 0.01) T cells proliferation under natural SEA, rSElK, and rSElQ treatment (Fig. 3). Moreover, addition of A549 into BALB/C mouse thymocytes induced more T cell proliferation than that of C57BL/6JT mouse (p < 0.01), which indicated a species-diversity in T cell activation. It might imply that TCR-Vβ subsets of T-lymphocytes differ in different mouse strains and various SEs had variable dependence on the Vβ subsets of TCR in the role of promoting lymphocyte proliferation.30
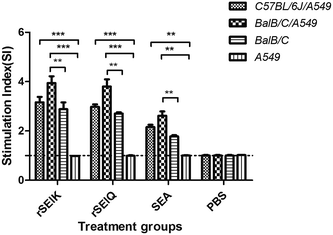 |
| | Fig. 3 SEs stimulation promotes T cells proliferation at presence of A549 cells. MTT assay was performed by using three kinds of cell samples subject to rSElK stimulation, rSElQ treatment and non-treatment as the negative control (X axis). Three columns separately represent the samples of C57BL/6J T-lymphocytes/A549 cells, BALB/C T-lymphocytes/A549 cells and BaB/C T-lymphocytes. Error bars represented the standard deviations, and statistical significance was determined by using Student's unpaired t-test comparing to PBS treatment. *P < 0.05; **P < 0.01; ***P < 0.001. | |
Effect of recombinant SEls on mouse viscera index (VI)
Mouse viscera index (VI) was also measured to reveal the superantigencity of rSElK and rSElQ in vivo in BALB/C mice. The two rSEls were injected into mice at a dose of 5 μg kg−1. After 72 h, spleen and thymus index was calculated to reflect the stimulating effect. As expected, rSElK, rSElQ induced evident viscera swelling compared to PBS control and showed a consistent result with the cell stimulation experiment in vitro (Fig. 4).
 |
| | Fig. 4 Low doses of recombinant SEs affect mouse spleen and thymus index. Fold change was calculated of mouse viscera (two columns, spleen and thymus) index after 72 h low-dose (5 μg kg−1) SEs (rSElK, rSElQ, natural SEA) stimulation compared with negative treatment (PBS). Error bars represent the standard deviations, and statistical significance was determined by using Student's unpaired t-test. *P < 0.05, **P < 0.01, ***P < 0.001 respectively for statistically significant, superior significant and the highest significant differences with respect to PBS treatment. | |
The ability of rSEls to induce cytokine production in vitro
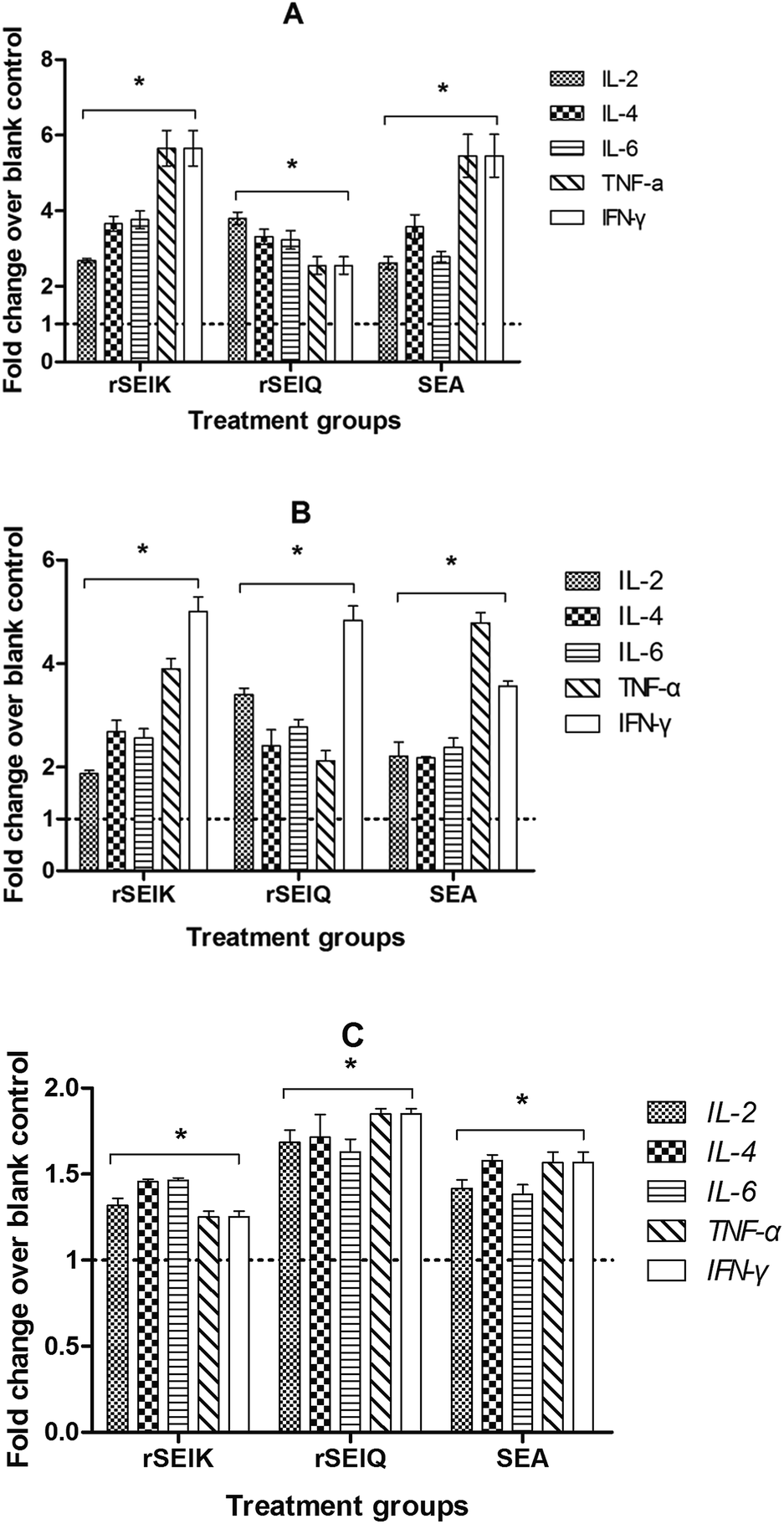
Bacterial enterotoxins, like staphylococcal enterotoxins, via superantigenic stimulation, lead to massive T cell proliferation and secretion of abnormally large amounts of proinflammatory cytokines. In order to consider the stimulating ability of SElK, Q to promote cytokines production, the transcriptional profile of a series of designated cytokines were evaluated, including IL-2, IL-4, IL-6, TNF-α and IFN-γ. We treated thymocytes from BALB/C or C57BL/6JT mouse with 40 ng mL−1 rSElK or rSElQ in presence or absence of A549 cells for 72 hours. Then the cytokine transcription was measured by relative quantitative RT-PCR shown in Fig. 5A–C. According to the results, rSElK and rSElQ were obvious to raise the level of cytokine transcription, especially in A549 cells co-incubation groups (Fig. 5A and B). And two different rSEls caused certain cytokine production to varying degrees. Specifically, the recombinant SEls existed a species-dependent stimulating activity in cytokine transcription, BALB/C mouse interacting with A549 cells triggered larger amount of cytokines expanding (Fig. 5A) than that of C57BL/6JT mouse (Fig. 5B). In addition, the protein production of these cytokines in the culture supernatants was also tested by ELISA. As shown in Fig. 6, rSElK and rSElQ both had a comparable way as SEA to incur the five cytokines expanding compared to PBS control.
 |
| | Fig. 5 The ability of rSEls to induce cytokine transcription in vitro. (A) The change rate of five cytokines transcription in BALB/C T-lymphocytes/A549 cells. (B) The effect of five cytokines transcription in C57BL/6J T-lymphocytes/A549 cells. (C) The result of five cytokines transcription in BALB/C T-lymphocytes. Total RNA from three kinds of cells treated by SEs (X axis) in vitro was applied for relative quantitative Real-Time PCR assay of five cytokines, IL-2, IL-4, IL-6, TNF-α, and IFN-γ. Fold change (Y axis) is cytokines change fold mentioned in the former. Error bars represent the standard deviations, and statistical significance was determined by using Student's unpaired t-test. *P < 0.05 for statistically significant differences with respect to PBS treatment. | |
 |
| | Fig. 6 The ability of rSEls to induce cytokine expression in vitro. Culture supernatants from SEs (three groups in X axis) and PBS stimulated BALB/C T cells were applied for ELISA assay of five cytokines, IL-2, IL-4, IL-6, TNF-α, IFN-γ (five columns). Y axis represents cytokines concentration. The changes of cytokine expressional level were discerned compared to PBS negative control. Error bars represent the standard deviations, and statistical significance was determined by using Student's unpaired t-test. *P < 0.05 for statistically significant differences with respect to PBS treatment. | |
rSEls vary in stimulating mouse cytokine generation in vivo
The immuno-stimulatory effect of two rSEls on the cytokine secretion was also examined in vivo. BALB/C mice were injected with 5 μg kg−1 amount of rSElK, rSElQ, SEA or PBS. At 72 hours post treatment, the thymocytes were isolated for total RNA extraction and the cytokine mRNA expression was determined by quantitative PCR. Meanwhile, an ELISA assay was carried out to measure level of the secreted cytokines in the serum. Consistent with the in vitro assays, rSElK and rSElQ induced a vast production of cytokines at both mRNA and protein levels by contrast to PBS control (Fig. 7A and B). As shown in Fig. 7A, rSElK had a superior effect on the transcription of cytokines such as IL-4, TNF-α, the level of which was 2 times higher than that of negative control. By contrast, treatment with PBS did not significantly increase the cytokine transcription. The same was true for the protein production of these cytokines in the sera as measured by ELISA.
 |
| | Fig. 7 rSEls vary in stimulating mouse cytokine generation in vivo. (A) Relative Real-Time fluorescent quantitation of five cytokine transcription by SEs treatment in vivo. Total RNA from SEs (three groups in X axis) stimulated BALB/C mouse thymus tissue was applied for relative quantitative Real-Time PCR of five cytokines, IL-2, IL-4, IL-6, TNF-α, IFN-γ (five columns). Fold change in Y axis represents cytokines change fold after treatment compared to the negative control. Fold change >1 indicates an increase in cytokines transcription after treatment, or is on the opposite. B. ELISA for five cytokines expression in vivo. Sera from SEs (three groups in X axis) and PBS stimulated BALB/C mouse were applied for ELISA assay of five cytokines, IL-2, IL-4, IL-6, TNF-α, IFN-γ (five columns). Y axis represents cytokines concentration. The changes of cytokine expressional level were discerned compared to PBS negative control. Error bars represent the standard deviations, and statistical significance was determined by using Student's unpaired t-test. *P < 0.05 for statistically significant differences with respect to PBS treatment. | |
Structure homology remodeling of rSEls
At a glance of distinct superantigenic activities related to SElK, SElQ, we tried to explore the reason of different function through homologous structure remodeling, typical SE structures consist of 5 alpha helices and 12 beta sheets (Fig. 8A). To search the molecular basis for the SElK and SElQ difference, we first analyzed the MHCII-binding sites on SEls (Fig. 8B). The structure of the putative MHC binding site on SElK and SElQ is also structurally homologous to that of SEI, and thus, they likely binds to MHC in a similar fashion as does the latter.41 As we know, SEs harbor two binding sites for MHC II molecules. One is the common sites, located in the N-terminal providing a weak affinity with MHCII molecules (e.g. SEB),42 while the other is zinc binding sites, located in the C-terminal having a strong affinity with MHCII molecules (e.g. SEH).43 The critical residues for zinc binding sites contain three amino acids: two histidines and an aspartic acid. This structure has a high affinity, approximately 100 times than that of N-terminal binding sites, making it the main sites for the SEs superantigenicity. It has been well documented that SEA contain binding sites for MHC II molecules, which consist of two histidine residues and an aspartic acid residue (asp227, his187, his225). Referring to our results, SElK and SElQ both possess a zinc-binding sites (Fig. 8B and Table 3).
 |
| | Fig. 8 Structure homology modeling of SElK and SElQ. (A) Three-dimensional representation of the two SE structures. The α-helices and β-stands were in rainbow colours from the N-(blue) to C-termini (red), respectively. α-Helices, β-stands as well as the N- and C-termini were labeled. (B) Zinc MHC-II binding sites of SElK, SElQ. Backbones, side chains of key residues (labelled) were shown as sticks. Red arrows in yellow circles (dashed lines) indicate Zinc MHC-II binding sites on key residues which were listed in Table 3. (C) α3–β8 loop on TCR binding sites of the two SEs. The loop between α3 and β8 was shown by C-backbones as sticks and colored according to the accessibility ranging in rainbow color from red (high) to blue (low). The key residues of each TCR binding site were labeled in pink dashed circles and detailed in Table 3. | |
Table 3 Function sites of SEs for MHC II and TCR binding
| Item |
MHCII |
TCR: α3–β8 loop |
| Zinc binding key amino acids |
Amino acid residues |
Key amino acids |
| SElK |
His169, His208, Asp210 |
26 |
142His, 158Tyr |
| SElQ |
His207, His245, Asp247 |
26 |
188Glu, 191Tyr |
Discussion
SEs have risen a wide range of concerns in human health. Existing epidemiological survey reveals a high prevalence rate (about 95.8% staphylococcal isolates harbored more than 2 SE/SEl genes) in different staphylococcal species (our unpublished data, not shown here). Specifically, Becker conducted a survey of the superantigen profile of 429 S. aureus from human blood samples or nasal discharges, and revealed a high detection rate of SEI and SEQ (55%),44 which arose our interest to have a deep research into new SEl types like SElK and SElQ. Meanwhile, amounting existing evidence, little is known about the superantigenic activity of SElK, SElQ and their role in food issues involved in public health. Hence we established a set of experiments in mice to evaluate the immune effect of rSElK and rSElQ. First, we measured viscera index of mouse thymus or spleen and conducted T cells proliferation by rSElK, rSElQ treatment compared with the SEA. It indicated a strong stimulating activity for both SEls to trigger T-lymphocyte activation and propagation. Meanwhile, relative quantitative PCR and ELISA analysis of inflammatory cytokines separately revealed a mass of cytokines transcription and expression in post treating samples. In some degree, those attempts partially demonstrated superantigenicity of the two recombinant SEls.
Superantigens cross bridge TCRs with MHC II molecules on APCs in a relatively nonspecific manner, inducing highly significant proliferation of T cells and activation of APCs such as macrophages.45–47 Although SEs can promote T-lymphocytes proliferation without APC cells, it can function well with APC participation. IFN-γ was an inducer for MHCII molecules expression on A549 cells, A549 lung carcinoma cells co-incubated with IFN-γ can promote vast amounts of MHCII molecules to incur more T-lymphocytes proliferation than T cells culture alone. It might be associated with MHCII-mediated signal transduction mechanisms.11 At the presence of MHCII molecules, it is capable of activating tyrosine kinase and membrane phosphoinositide, which eventually leads to expression of inflammatory cytokines and further results in an abundant release of cytokines, followed with IFN-γ, IL-2 stimulating further differentiation and proliferation of T-lymphocytes.11,22
Previous researches have demonstrated that the SEs superantigens interact with TCR in three modes: first, SEs specifically recognize and bind to certain amino acid residues in CDR2, FR3 regions of TCR, such as TSST.45,48 Second, SEs specifically recognize spatial structure of CDR2, FR3 rather than amino sequence of Vβ, such as SEB, SEC.46,49,50 Third, α3–β8 loop of SEs specifically bonds FR3, FR4 regions of TCR, such as SElK, SEI.50 α3–β8 loop is a hypervariable region which has been found in both SElK and SElQ (Table 3). This loop binds to TCR via two different modes, either directly recognizing certain residues or binding to spatial structure of Vβ, it is critical for the specificity of the interaction of the superantigens with their respective Vβ-TCRs.47 For SEA, three key amino acid residues with 14 aa length of the α3–β8 loop are implicated in TCR binding, in which are two Asp residues and one Ser (Ser172, Asp173, Asp175) forming a strong hydrogen bond with the Vβ amino acids. In this study, the two proteins were at the similar length of α3–β8 loop with 26aa amino acids but possessed different residues. A long length of amino acids (26aa) has a good flexibility and mutable structure for Vβ bonding. For SElK, there are 2 residues (His142, Tyr158) implicated in TCR binding whilst SElQ with Glu188 and Tyr191 (Fig. 8C). The Glu residue with a good hydrophilicity can be exposed on molecular surface and is capable of forming hydrogen bonds along with a –OH on phenyl loop. This provides easier accessibility to affine with TCR. In contrast, the predicted residue (His) that is exposed on the surface is only capable of forming intra-molecular hydrogen bonds, making it a lower affinity, and the numerous residues within these SEs–TCRVβ complex structure interfaces are likely to contribute significantly to both binding and specificity.
In summary, the SElK and SElQ can form signaling complexes with MHC and TCR Vβ molecules. The MHC–SEs–TCR ternary complex have significant functional consequences and can activate SEs-specific T cell signaling pathway to boost cell proliferation and cause the cytokine storm. Thus, the enhanced activity of SEIK compared to SEIQ and SEA, their structural difference, binding efficiency, variations in their MHC-II binding pocket etc., all those are contributed to their superantigenicity and functions. The structural difference (e.g. the critical amino acids of the α3–β8 loop) might partially explain the diverse immune-stimulatory activity of SElK and SElQ. In accordance with the diverse proliferation scale of T cells from different mouse series, it should also be kept in mind that SEls only recognize certain subtypes of TCR Vβ. The Vβ subtypes in the bracket are recognized by SElK (5.1, 5.2, 6.7), SElQ (2.1, 5.1, 6.7, 21.3) of human. Proliferation of BALB/C T-lymphocytes was superior to C57BL/6J mouse, which implied that BALB/C mouse with Vβ subtypes of certain types could be recognized by these two SEs just missing in C57BL/6J mice or with more SEs recognition sites in BALB/C T cells than another. Some specific TCR Vβ domain have been shown to be overrepresented in some species and patients with Crohn's disease, a severe inflammatory bowel syndrome.51
Conflict of interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author contributions
Conceived and designed the experiments: Jinhai Huang. Performed the experiments: Yihe Xia, Liu Yang, Zhixuan Liang, Xiumei Li, Xianzhi He. Analyzed the data: Yihe Xia, Hui Deng. Contributed reagents/materials/analysis tools: Jinhai Huang. Wrote the paper: Jinhai Huang.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (no. 31272540) and the National high technology research and development program of China (863 program, no. 2012AA101605).
References
- T. J. Foster, Staphylococci and staphylococcal infections, Expert Rev. Anti-Infect. Ther., 2010, 8, 1337–1338 CrossRef PubMed.
- B. L. Kotzin, D. Y. Leung, J. Kappler and P. Marrack, Superantigens and their potential role in human disease, Adv. Immunol., 1993, 54, 99–166 CAS.
- S. Holtfreter and B. Broker, Staphylococcal superantigens: do they play a role in sepsis, Arch. Immunol. Ther. Exp., 2005, 53, 13–27 CAS.
- J. K. McCormick, J. M. Yarwood and P. M. Schlievert, Toxic shock syndrome and bacterial superantigens: an update, Annu. Rev. Microbiol., 2001, 55, 77–104 CrossRef CAS PubMed.
- N. Balaban and A. Rasooly, Staphylococcal enterotoxins, Int. J. Food Microbiol., 2000, 61, 1–10 CrossRef CAS.
- M. S. Bergdoll, R. N. Robbins, K. Weiss, C. R. Borja, Y. Huang and F. S. Chu, The staphylococcal enterotoxins: similarities, Contrib. Microbiol. Immunol., 1973, 1, 390–396 CAS.
- P. Marrack and J. Kappler, The staphylococcal enterotoxins and their relatives, Science, 1990, 248, 705–711 CAS.
- M. D. Baker and K. R. Acharya, Superantigens: structure–function relationships, IJMM, Int. J. Med. Microbiol., 2004, 293, 529–537 CrossRef CAS PubMed.
- Y.-W. Choi, B. Kotzin, L. Herron, J. Callahan, P. Marrack and J. Kappler, Interaction of Staphylococcus aureus toxin“ superantigens” with human T cells, Proc. Natl. Acad. Sci. U. S. A., 1989, 86, 8941–8945 CrossRef CAS.
- J. Kappler, B. Kotzin, L. Herron, E. W. Gelfand, R. D. Bigler, A. Boylston, S. Carrel, D. N. Posnett, Y. Choi and P. Marrack, V beta-specific stimulation of human T cells by staphylococcal toxins, Science, 1989, 244, 811–813 CAS.
- J. Yagi, J. Baron, S. Buxser and C. Janeway, Bacterial proteins that mediate the association of a defined subset of T cell receptor: CD4 complexes with class II MHC, J. Immunol., 1990, 144, 892–901 CAS.
- M. M. Dinges, P. M. Orwin and P. M. Schlievert, Exotoxins of Staphylococcus aureus, Clin. Microbiol. Rev., 2000, 13, 16–34 CrossRef CAS PubMed.
- C. W. Ronson, B. T. Nixon and F. M. Ausubel, Conserved domains in bacterial regulatory proteins that respond to environmental stimuli, Cell, 1987, 49, 579–581 CrossRef CAS.
- S. H. Munson, M. T. Tremaine, M. J. Betley and R. A. Welch, Identification and characterization of staphylococcal enterotoxin types G and I from Staphylococcus aureus, Infect. Immun., 1998, 66, 3337–3348 CAS.
- H. Kuang, W. Wang, L. Xu, W. Ma, L. Liu, L. Wang and C. Xu, Monoclonal Antibody-Based Sandwich ELISA for the Detection of Staphylococcal Enterotoxin A, Int. J. Environ. Res. Public Health, 2013, 10, 1598–1608 CrossRef CAS PubMed.
- Q. Jiao, H. Wang, Z. Hu, Y. Zhuang, W. Yang, M. Li, X. Yu, J. Liang, Y. Guo and H. Zhang, Lidocaine inhibits staphylococcal enterotoxin-stimulated activation of peripheral blood mononuclear cells from patients with atopic dermatitis, Arch. Dermatol. Res., 2013, 1–8 Search PubMed.
- Y. Le Loir, F. Baron and M. Gautier, Staphylococcus aureus and food poisoning, Genet. Mol. Res., 2003, 2, 63–76 Search PubMed.
- A. Aydin, M. Sudagidan and K. Muratoglu, Prevalence of staphylococcal enterotoxins, toxin genes and genetic-relatedness of foodborne Staphylococcus aureus strains isolated in the Marmara region of Turkey, Int. J. Food Microbiol., 2011, 148, 99–106 CrossRef CAS PubMed.
- P. M. Orwin, D. Y. Leung, H. L. Donahue, R. P. Novick and P.
M. Schlievert, Biochemical and biological properties of staphylococcal enterotoxin K, Infect. Immun., 2001, 69, 360–366 CrossRef CAS PubMed.
- P. M. Orwin, D. Y. Leung, T. J. Tripp, G. A. Bohach, C. A. Earhart, D. H. Ohlendorf and P. M. Schlievert, Characterization of a novel staphylococcal enterotoxin-like superantigen, a member of the group V subfamily of pyrogenic toxins, Biochemistry, 2002, 41, 14033–14040 CrossRef CAS PubMed.
- M. S. Bergdoll, Enterotoxins, Staphylococci Staphylococcal Infect., 1983, 2, 559–598 CAS.
- G. A. Bohach, D. J. Fast, R. D. Nelson and P. M. Schlievert, Staphylococcal and streptococcal pyrogenic toxins involved in toxic shock syndrome and related illnesses, Crit. Rev. Microbiol., 1990, 17, 251–272 CrossRef CAS PubMed.
- A. J. Brosnahan and P. M. Schlievert, Gram-positive bacterial superantigen outside-in signaling causes toxic shock syndrome, FEBS J., 2011, 278, 4649–4667 CrossRef CAS PubMed.
- J. L. Aguilar, A. K. Varshney, X. Wang, L. Stanford, M. Scharff and B. C. Fries, Detection and measurement of staphylococcal enterotoxin-like K (SEl-K) secretion by Staphylococcus aureus clinical isolates, J. Clin. Microbiol., 2014, 52, 2536–2543 CrossRef CAS PubMed.
- L. Zeng, J. Huang, Y. Liu, S. Zhuang and Z. Xue, Bioactivity of the recombinant staphylococcal enterotoxin Q and its structure relationship analysis, Acta Agric. Boreali-Occident. Sin., 2010, 25, 80–84 Search PubMed.
- Y. Q. Pan, D. Ding, D. X. Li and S. Q. Chen, Expression and bioactivity analysis of staphylococcal enterotoxin M and N, Protein Expression Purif., 2007, 56, 286–292 CrossRef CAS PubMed.
- J. Sambrook and D. W. Russell, Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y, 2001 Search PubMed.
- Y.-C. Su and A. Wong, Identification and purification of a new staphylococcal enterotoxin, H, Appl. Environ. Microbiol., 1995, 61, 1438–1443 CAS.
- Q. Xue, Y. B. Ying, Y. Q. Pan, D. X. Li, H. Y. Sun and S. Q. Chen, Expression and bioactivity analysis of staphylococcal enterotoxin C2, Yaoxue Xuebao, 2006, 41, 406–411 CAS.
- T. Mosmann, Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays, J. Immunol. Methods, 1983, 65, 55–63 CrossRef CAS.
- S. Swaminathan, W. Furey, J. Pletcher and M. Sax, Residues defining V beta specificity in staphylococcal enterotoxins, Nat. Struct. Biol., 1995, 2, 680–686 CrossRef CAS PubMed.
- E. J. Sundberg, M. W. Sawicki, S. Southwood, P. S. Andersen, A. Sette and R. A. Mariuzza, Minor structural changes in a mutated human melanoma antigen correspond to dramatically enhanced stimulation of a CD4+ tumor-infiltrating lymphocyte line, J. Mol. Biol., 2002, 319, 449–461 CrossRef CAS.
- M. J. Cook, The anatomy of the laboratory mouse, 1965 Search PubMed.
- M. J. Lay and C. T. Wittwer, Real-Time fluorescence genotyping of factor V Leiden during rapid-cycle PCR, Clin. Chem., 1997, 43, 2262–2267 CAS.
- U. Reischl, H.-J. Linde, M. Metz, B. Leppmeier and N. Lehn, Rapid identification of methicillin-resistant Staphylococcus aureus and simultaneous species confirmation using Real-Time fluorescence PCR, J. Clin. Microbiol., 2000, 38, 2429–2433 CAS.
- K. J. Livak and T. D. Schmittgen, Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔC Method, Methods, 2001, 25, 402–408 CrossRef CAS PubMed.
- E. Schad, I. Zaitseva, V. Zaitsev, M. Dohlsten, T. Kalland, P. Schlievert, D. Ohlendorf and L. Svensson, Crystal structure of the superantigen staphylococcal enterotoxin type A, EMBO J., 1995, 14, 3292 CAS.
- K. Mehindate, J. Thibodeau, M. Dohlsten, T. Kalland, R.-P. Sekaly and W. Mourad, Cross-linking of major histocompatibility complex class II molecules by staphylococcal enterotoxin A superantigen is a requirement for inflammatory cytokine gene expression, J. Exp. Med., 1995, 182, 1573–1577 CrossRef CAS.
- M. Langford, G. Stanton and H. Johnson, Biological effects of staphylococcal enterotoxin A on human peripheral lymphocytes, Infect. Immun., 1978, 22, 62–68 CAS.
- D. A. Hildeman, Y. Zhu, T. C. Mitchell, J. Kappler and P. Marrack, Molecular mechanisms of activated T cell death in vivo, Curr. Opin. Immunol., 2002, 14, 354–359 CrossRef CAS.
- M. M. Fernandez, R. Guan, C. P. Swaminathan, E. L. Malchiodi and R. A. Mariuzza, Crystal structure of staphylococcal enterotoxin I (SEI) in complex with a human major histocompatibility complex class II molecule, J. Biol. Chem., 2006, 281, 25356–25364 CrossRef CAS PubMed.
- T. S. Jardetzky, J. H. Brown, J. C. Gorga, L. J. Stern, R. G. Urban, Y.-i. Chi, C. Stauffacher, J. L. Strominger and D. C. Wiley, Three-dimensional structure of a human class II histocompatibility molecule complexed with superantigen, Nature, 1994, 368(6473), 711–718 CrossRef CAS PubMed.
- M. Håkansson, K. Petersson, H. Nilsson, G. Forsberg, P. Björk, P. Antonsson and L. Svensson, The crystal structure of staphylococcal enterotoxin H: implications for binding properties to MHC class II and TCR molecules, J. Mol. Biol., 2000, 302, 527–537 CrossRef PubMed.
- K. Becker, A. W. Friedrich, G. Lubritz, M. Weilert, G. Peters and C. von Eiff, Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens, J. Clin. Microbiol., 2003, 41, 1434–1439 CrossRef CAS.
- B. Moza, R. A. Buonpane, P. Zhu, C. A. Herfst, A. N.-U. Rahman, J. K. McCormick, D. M. Kranz and E. J. Sundberg, Long-range cooperative binding effects in a T cell receptor variable domain, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 9867–9872 CrossRef CAS PubMed.
- H. Li, A. Llera, D. Tsuchiya, L. Leder, X. Ysern, P. M. Schlievert, K. Karjalainen and R. A. Mariuzza, Three-dimensional structure of the complex between a T cell receptor β chain and the superantigen staphylococcal enterotoxin B, Immunity, 1998, 9, 807–816 CrossRef CAS.
- S. Günther, A. K. Varma, B. Moza, K. J. Kasper, A. W. Wyatt, P. Zhu, A. Rahman, Y. Li, R. A. Mariuzza and J. K. McCormick, A novel loop domain in superantigens extends their T cell receptor recognition site, J. Mol. Biol., 2007, 371, 210–221 CrossRef PubMed.
- B. Moza, A. K. Varma, R. A. Buonpane, P. Zhu, C. A. Herfst, M. J. Nicholson, A.-K. Wilbuer, N. P. Seth, K. W. Wucherpfennig and J. K. McCormick, Structural basis of T-cell specificity and activation by the bacterial superantigen TSST-1, EMBO J., 2007, 26, 1187–1197 CrossRef CAS PubMed.
- B. A. Fields, E. L. Malchiodi, H. Li, X. Ysern, C. V. Stauffacher, P. M. Schlievert, K. Karjalainen and R. A. Mariuzza, Crystal structure of a T-cell receptor β-chain complexed with a superantigen, Nature, 1999, 384(6605), 188–192 CrossRef PubMed.
- Y. Li, H. Li, N. Dimasi, J. K. McCormick, R. Martin, P. Schuck, P. M. Schlievert and R. A. Mariuzza, Crystal structure of a superantigen bound to the high-affinity, zinc-dependent site on MHC class II, Immunity, 2001, 14, 93–104 CrossRef CAS.
- T. P. Prindiville, M. C. Cantrell, T. Matsumoto, W. R. Brown, A. A. Ansari, B.
L. Kotzin and M. E. Gershwin, Analysis of function, specificity and T cell receptor expression of cloned mucosal T cell lines in Crohn's disease, J. Autoimmun., 1996, 9, 193–204 CrossRef CAS.
Footnote |
| † These author contributed equally to this work. |
|
| This journal is © The Royal Society of Chemistry 2015 |
Click here to see how this site uses Cookies. View our privacy policy here.