
Discovery of new targets of phenolic acids in danshen using a label-free cell phenotypic assay†
Jixia Wanga,
Tao Houb,
Lai Weia,
Liying Shib,
Jian Hea,
Nan Zhoud,
Guangwei Suna,
Xiuli Zhang*ac and
Xinmiao Lianga
aKey Lab of Separation Science for Analytical Chemistry, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian, China. E-mail: zhangxiuli@dicp.ac.cn; Fax: +86 411 84379539; Tel: +86 411 84379519
bInstitute of Materia Medica, Dalian University, Dalian, China
cCo-innovation Center of Neuroregeneration, Nantong University, Nantong, 226019, China
dDepartment of Oral Pathology, College of Stomatology, Dalian Medical University, Dalian, China
First published on 26th February 2015
Abstract
Traditional Chinese medicines (TCMs) have been used in the clinic for thousands of years. Their reliable therapeutic efficacies are closely related to their multi-target mechanisms of action (MOAs). Discovery of these targets is important for understanding their clinical features. Danshen is the dried root of Salvia miltiorrhiza, a Traditional Chinese Medicine (TCM) used for the treatment of cardiovascular and cerebrovascular diseases. Although its clinical features are well recognized, the targets of its active constituents are poorly understood. Here, a label-free cell phenotypic assay was used to investigate the potential targets of the phenolic acids in danshen. Pharmacological profiling of 10 known phenolic acids in danshen using HT-29 and A431 cells revealed that lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and danshensu all displayed agonistic activity towards GPR35; however, salvianolic acid A and salvianolic acid C at high doses were also active in altering intracellular Ca2+ via another unknown target. Since GPR35 has been implicated in inflammation and cardiovascular diseases, the discovery of GPR35 as one target of the phenolic acids in danshen was useful for elucidating their mechanisms of action in the treatment of these diseases. This study also highlights the potential of label-free cell phenotypic assays for discovering multiple targets of TCMs.
1. Introduction
Traditional Chinese Medicines (TCMs) are attracting considerable attention owing to their reliable clinical efficacy, thus being important resources for lead compounds or drugs.1,2 Most of them provide therapeutic effect by interacting with multiple targets. For instance, paclitaxel (anticancer), reserpine (antihypertensive), artemisinin (antimalarial) and rapamycin (immunosuppression) act on at least two targets. Studying these mechanisms of action (MOAs) is important for understanding their clinical features. Target-based approaches that screen compounds against a single target protein are not effective for discovering multiple targets of compounds.3 However, phenotype-based approaches can provide an unbiased way to find active compounds, and are useful for identifying multiple targets or pathways.4As one of a new generation of phenotype-based assay techniques, the label-free cell phenotypic assay has become an attractive approach for determining the target engagement of drugs.5–7 This assay uses a label-free resonant waveguide grating (RWG) biosensor to convert drug-induced dynamic redistribution of cellular constituents in cells into an integrated and kinetic response, termed dynamic mass redistribution (DMR).14 The DMR is recorded as a shift in resonant wavelength in picometers (pm) and represents a cell phenotypic response covering a wide range of targets/pathways.6–9 Furthermore, due to its non-invasiveness, the DMR assay can be performed in multiple formats and permits intervention with probe molecules, thus enabling mechanistic elucidation of drug pharmacology.10–12
Danshen, the dried root of Salvia miltiorrhiza, is a Traditional Chinese Medicine (TCM) widely used in China for the treatment of cardiovascular and cerebrovascular diseases.13,14 The danshen dripping pill is currently in phase III clinical trials with a great hope to be the first Food and Drug Administration (FDA) approved TCM. Danshen extract contains two main types of ingredients, water-soluble phenolic acids and lipophilic diterpenoid quinines. Until now, more than 100 compounds have been isolated,15 which include about 30 phenolic acid compounds. According to the number of caffeic acid units, these phenolic acids can be classified into five subgroups: monomers, dimers, trimers, tetramers and others.14 These compounds occur in the danshen extract in various concentrations. Based on fingerprinting analysis of the danshen extract,16 caffeic acid, danshensu, rosmarinic acid and salvianolic acid B exist in all the studied species; salvianolic acid B and rosmarinic acid are relatively rich in the extract. In terms of danshen preparations,17 the tablets and capsules contain both water-soluble phenolic acids and lipophilic diterpenoid quinines, while the dropping pills and injections contain mainly phenolic acids. Among these phenolic acids, the contents of danshensu, protocatechuic aldehyde and salvianolic acid A were higher than those in other solid samples. Salvianolic acid B is the predominant compound in the crude extract, but decreases in the dropping pills and injections. Given that a water decoction and an injection agent are the common forms of danshen in the clinic, the water-soluble phenolic acids are believed to be responsible for its biological activities. These water-soluble components have been shown to have activities as antioxidant, anti-ischemia reperfusion, anti-hypertension, antiplatelet aggregation, anti-inflammation and anti-fibrosis agents.18 Determining target engagement represents an important step towards understanding the pharmacological activities of the water-soluble phenolic acids in danshen, and thus the clinical features of danshen products.19 Recently, in silico analysis and experimental validation have demonstrated that salvianolic acid A exhibits an anti-inflammatory effect via the induction of heme oxygenase 1 through the p38 MAPK pathway in lipopolysaccharide (LPS)-stimulated RAW264.7 cells.20 Salvianolic acid B may inhibit angiotensin converting enzyme (ACE) and thus attenuate angiotensin I-induced vasoconstriction, which might partially account for its anti-hypertension effect.21 Salvianolic acid B may also prevent cardiac remodeling by inhibiting matrix metalloproteinase-9.22 Furthermore, salvianolic acid A, salvianolic acid B and caffeic acid can all inhibit protein–protein interactions via the SH2 domains of the Src-family kinases, Src and Lck.23 These findings suggest that these phenolic acids may interact with multiple targets rather than a single target. Therefore, further discovery of new targets of phenolic acids in danshen is of importance to understand and elucidate their clinical features.
Here we use a label-free cell phenotypic assay to discover new targets of phenolic acids from danshen with a hope to elucidate its in vivo therapeutic effects.
2. Experimental
2.1. Materials
Lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C, danshensu, caffeic acid, isoferulic acid, carnosol and protocatechuic aldehyde were purchased from Shanghai Yuanye Bio-technology Co., Ltd. (China). Salvianolic acid D was gifted from Prof. Jingbo Zhu (Dalian Polytechnic University). Their structures are shown in Fig. 1. Acetylcholine chloride, bradykinin, cyclopiazonic acid, histamine, kynurenic acid, thapsigargin, zaprinast and SPB05142 (CID2745687) were obtained from Sigma Chemical Co. (St Louis, MO, USA). ML145 was obtained from Tocris Bioscience Co. (St. Louis, MO, USA). Rabbit polyclonal β-actin antibody, rabbit polyclonal extracellular-signal regulated kinase 1/2 antibody (anti-ERK1/2) and rabbit polyclonal anti-GPR35 were obtained from Proteintech (Wuhan, China). Rabbit polyclonal anti-phosphorylated extracellular-signal-regulated kinase 1/2 (Thr202/Tyr20) was from Bioworld Technology, Co. Ltd. (Naijing, China). Epic® 384-well biosensor cell culture compatible microplates were obtained from Corning Incorporated (Corning, NY, USA). Human colorectal adenocarcinoma HT-29 and human epidermoid carcinoma A431 cell lines were obtained from the Cell Bank of Shanghai Institute of Cell Biology, Chinese Academy of Sciences.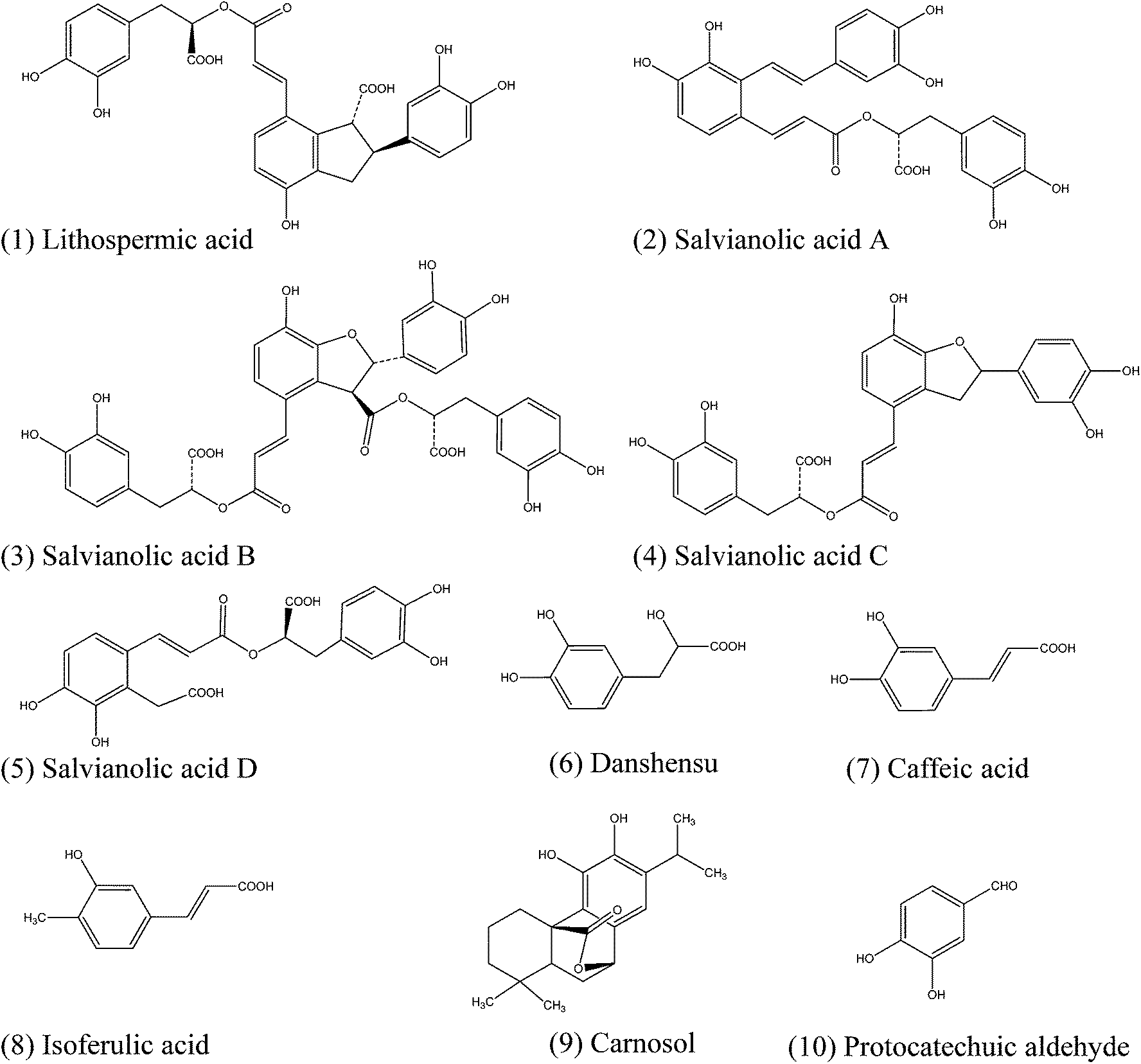 | ||
| Fig. 1 Chemical structures of 10 known phenolic acids in danshen. | ||
Acetylcholine chloride, cyclopiazonic acid, danshensu, lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and thapsigargin were stocked at 100 mM, while all other compounds were stocked at 10 mM. With the exception of acetylcholine, which was prepared in water, all other compounds were dissolved in 100% dimethyl sulfoxide (DMSO). All compounds were freshly diluted with the assay buffer (1× Hank’s balanced salt solution (HBSS) buffer, 10 mM Hepes, pH 7.2) to the assayed concentrations.
2.2. Cell culture
HT-29 cells were cultured using McCoy’s 5A Medium (#DY0324S0414L, Sango Biotech, Shanghai, China) with 10% fetal bovine serum (#10099141, Gibco, Life Technologies), 50 μg mL−1 penicillin and 100 μg mL−1 streptomycin in a humidified 37 °C/5% CO2 incubator. A431 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (#SH30022.01B, Thermo Scientific HyClone) supplemented with 10% fetal bovine serum, 50 μg mL−1 penicillin and 100 μg mL−1 streptomycin at 37 °C under air/5% CO2.2.3. Dynamic mass redistribution assay
All DMR assays were performed using an Epic® BT system (Corning, NY, USA). Cells were directly seeded in Epic 384-well biosensor microplates with a seeding density of 32![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 and 25000 cells per well for HT-29 and A431 cells, respectively. After being cultured for ∼20 h, these cells formed a monolayer in the cell culture medium with a confluency of ∼95%. A431 cells were further starved for 24 h using serum-free medium. After culture, the cells were manually washed using the assay buffer and maintained with 30 μL of the assay buffer for 1 h in the Epic system.
000 and 25000 cells per well for HT-29 and A431 cells, respectively. After being cultured for ∼20 h, these cells formed a monolayer in the cell culture medium with a confluency of ∼95%. A431 cells were further starved for 24 h using serum-free medium. After culture, the cells were manually washed using the assay buffer and maintained with 30 μL of the assay buffer for 1 h in the Epic system.
For profiling compounds in HT-29 cells, a 2 min baseline was first established, followed by adding compounds and monitoring the compound-induced DMR signals for 1 h. Then a 2 min baseline was re-established, followed by adding 1 μM zaprinast or 16 μM acetylcholine and monitoring the DMR signals for another 1 h. Similarly, for profiling compounds in A431 cells, a 2 min baseline was first established, followed by compound addition and DMR recording for 1 h. A 2 min baseline was then re-established, followed by adding 16 nM bradykinin or 1 μM histamine and monitoring the DMR signals for 1 h.
For the DMR agonist assay, after establishment of a 2 min baseline, phenolic acids at varied doses were added individually and the DMR signals were recorded for 1 h. For the DMR desensitization assay, cells were pretreated with these compounds for 1 h, followed by recording a 2 min baseline, adding 1 μM zaprinast and monitoring the DMR signals for 1 h. For the DMR antagonist assay, HT-29 cells were pretreated with an antagonist, SPB05142 for 5 min or ML145 for 10 min, following by recording a 2 min baseline, adding a ligand at its indicated concentration (EC20, EC50 or EC80) and monitoring the DMR signals for 1 h.
To examine compounds for altering intracellular Ca2+, a two-step assay was also used. Different doses of salvianolic acid A, salvianolic acid C, thapsigargin and cyclopiazonic acid were first assayed, followed by recording the DMR signal induced by cyclopiazonic acid at 16 μM for 1 h.
2.4. Western blot
After the cells were treated with compounds for 15 min, whole cell lysates were harvested using lysis buffer (KeyGEN BioTECH, Naijing, China). For each group, 20 μg total protein was electrophoresed through a 12% (w/v) acrylamide gel and electroblotted onto polyvinylidene difluoride (PVDF) membranes. Antibodies were diluted as follows: β-actin (1:300), ERK1/2 (1:300), p-ERK1/2Thr202/Tyr204 (1:1000) and GPR35 (1:500). Detection was performed using a GTVTM III Detection System/Mo&Rb (GeneTech, Shanghai, China).
2.5. Data analysis
All DMR data were acquired using Epic Imager software (Corning, NY, USA) and processed using Imager Beta 3.7 (Corning), Microsoft Excel 2010 and GraphPad Prism 6.02 (GraphPad Sofware Inc., San Diego, CA, USA). All DMR signals were background corrected. All EC50 or IC50 values described were calculated based on the maximal amplitude of the DMR signal within 40 min post-stimulation. All DMR data were from two independent measurements, each in duplicate (n = 4).3. Results and discussion
3.1. Profiling of phenolic acids from danshen
We first profiled the activity of 10 phenolic acids in danshen using a RWG biosensor-enabled label-free cell phenotypic assay in HT-29 and A431 cells. The concentration of each compound for profiling was 10 μM. These phenolic acids are known to be present in danshen (Fig. 1). A two-step DMR assay was performed. The first step was a DMR agonist assay to characterize all compounds for their agonist activities in HT-29 and A431 cells. The second step was a DMR desensitization assay to examine the ability of each compound to desensitize the cellular response induced by a panel of agonist probe molecules, each at its saturating dose. Since HT-29 cells endogenously express GPR3524 and muscarinic M3 receptor,12,25 zaprinast26 and acetylcholine12 were chosen as their agonist probes, respectively. Similarly, A431 endogenously express bradykinin B2 receptor25,26 and histamine H1 receptor,27 so bradykinin and histamine were chosen as agonist probes, respectively. Based on the EC100 reported in the literature, the doses of the probe agonists examined were set at 1 μM, 16 μM, 16 nM and 1 μM for zaprinast, acetylcholine, bradykinin and histamine, respectively. Additionally, the expression level of GPR35 in HT-29 and A431 cells was examined using western blot and DMR assays (Fig. S1†). The results indicated that the expression level of GPR35 in HT-29 cells was much higher than that in A431 cells.
The results showed that in HT-29 cells salvianolic acid A and salvianolic acid C triggered a large DMR, while lithospermic acid, salvianolic acid B and danshensu led to a relatively small DMR, and all of the other five compounds resulted in negligible DMR (Fig. 2a). Interestingly, the five active compounds, including salvianolic acid A, salvianolic acid B, salvianolic acid C, lithospermic acid and danshensu, all attenuated the zaprinast-induced signal (Fig. 2b). Furthermore, the ability of salvianolic acid A and salvianolic acid C to attenuate the zaprinast-induced signal was greater than that of lithospermic acid, salvianolic acid B and danshensu. However, all compounds had no or little effect on the acetylcholine-induced signal (Fig. 2c). Combining the agonistic activity with the desensitization pattern suggests that lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and danshensu may have agonistic activity against GPR35.
 | ||
| Fig. 2 A label-free cell phenotypic profiling technique to identify the potential polypharmacology of phenolic acid compounds in HT-29 cells (a–c) and A431 cells (d–f). (a) The DMR amplitudes at 30 min post-stimulation of 10 μM compounds as a function of compounds in HT-29 cells. (b) The DMR response of 1 μM zaprinast after the cells were pre-stimulated with each compound for 1 h. (c) The DMR response of 16 μM acetylcholine after the cells were pre-stimulated with each compound for 1 h. (d) The DMR amplitudes of 10 μM compounds as a function of compounds in A431 cells. (e) The DMR response of 16 nM bradykinin after the cells were pre-stimulated with each compound for 1 h. (f) The DMR response of 1 μM histamine after the cells were pre-stimulated with each compound for 1 h. The compounds are lithospermic acid (1), salvianolic acid A (2), salvianolic acid B (3), salvianolic acid C (4), salvianolic acid D (5), danshensu (6), caffeic acid (7), isoferulic acid (8), carnosol (9), protocatechuic aldehyde (10) and control (11). All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
Different label-free pharmacological patterns were observed in A431 cells. The DMR responses at 30 min post-stimulation showed that salvianolic acid A and salvianolic acid C triggered a relatively strong DMR signal, while lithospermic acid and salvianolic acid B led to a small DMR signal, and other compounds were inactive (Fig. 2d). Furthermore, both salvianolic acid A and salvianolic acid C attenuated the bradykinin- and histamine-induced DMR signals (Fig. 2e and f, respectively). Given that in A431 cells there is little GPR35 expressed (Fig. S1†), histamine activates the H1 receptor triggering Gq-signaling,5 and bradykinin activates the B2 receptor triggering both Gs- and Gq-pathways,27 these results suggest that salvianolic acid A and salvianolic acid C may alter intracellular Ca2+ via another unknown target.
3.2. Activity of phenolic acids against GPR35
We next examined the potential agonistic activity of lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and danshensu against GPR35 using three different DMR assays. The DMR agonist assay was employed to determine their agonist activities. The DMR desensitization assay and DMR antagonist assay were used to examine the specificity of these five compounds for activating GPR35.First, the DMR agonist assay showed that lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and danshensu in the concentration range of 0.03–250 μM all triggered dose-dependent positive DMR signals, but with different amplitudes (Fig. 3). According to the DMR characteristics, these five compounds can be divided into two groups. The first group consists of salvianolic acid A and salvianolic acid C, both triggering a quite large DMR with almost identical characteristics (Fig. 3a). The second group consists of lithospermic acid, salvianolic acid B and danshensu, all triggering a moderate DMR (Fig. 3b). Except for salvianolic acid B and danshensu, whose DMR signals seem not to reach saturation at the maximal concentration of 250 μM examined, the dose responses of salvianolic acid A, salvianolic acid C and lithospermic acid were best fitted with a single-phase non-linear regression (Fig. 3c and d). The EC50 values of these compounds are shown in Table 1. Furthermore, the maximal DMR of lithospermic acid, salvianolic acid B and danshensu all shared similarity with those of zaprinast and kynurenic acid (Fig. 4). Zaprinast26 and kynurenic acid28 are two known GPR35 agonists and are often used as probe agonists for GPR35. Of note, salvianolic acid A or salvianolic acid C at 16 μM gave rise to a DMR also similar to that of zaprinast, but at high doses triggered a DMR that is much greater than the maximal DMR of zaprinast, suggesting that both compounds may also activate another unknown target or pathway, besides GPR35.
 | ||
| Fig. 3 DMR dose responses of compounds in HT-29 cells. (a and b) Real time DMR signals of salvianolic acid A (a) and danshensu (b); (c and d) DMR amplitudes as a function of compound doses: salvianolic acid A and salvianolic acid C (c), salvianolic acid B, lithospermic acid and danshensu (d). All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
| Compounds | EC50 (μM) | IC50a (μM) | IC50b (μM) | IC50c (μM) |
|---|---|---|---|---|
| a IC50 to desensitize the DMR of 1 μM zaprinast in HT-29 cells.b IC50 of SPB05142 to block the DMR of a phenolic acid compound at doses of 125 μM, 100 μM, 100 μM, 15 μM and 4 μM for lithospermic acid, salvianolic acid B, danshensu, salvianolic acid A and salvianolic acid C, respectively. The HT-29 cells were pre-treated with the antagonist for 5 min.c IC50 of ML145 to block the DMR of compounds at the same doses as above. The HT-29 cells were pre-treated with the antagonist for 10 min. | ||||
| Lithospermic acid | 37.63 ± 13.83 | 109.04 ± 23.79 | 2.29 ± 0.52 | 0.47 ± 0.07 |
| Salvianolic acid A | 37.69 ± 3.77 | 3.48 ± 0.38 | 2.52 ± 0.48 | 0.15 ± 0.10 |
| Salvianolic acid B | Weak | 72.74 ± 22.96 | 0.71 ± 0.42 | 1.48 ± 0.22 |
| Salvianolic acid C | 18.83 ± 1.11 | 2.70 ± 0.31 | 1.57 ± 0.64 | 0.24 ± 0.03 |
| Danshensu | Weak | 30.42 ± 2.25 | 2.98 ± 1.35 | 0.25 ± 0.02 |
 | ||
| Fig. 4 DMR dose responses of probes in HT-29 cells. (a and b) Real time DMR signals of zaprinast (a) and kynurenic acid (b); (c) DMR amplitudes as a function of compound doses. All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
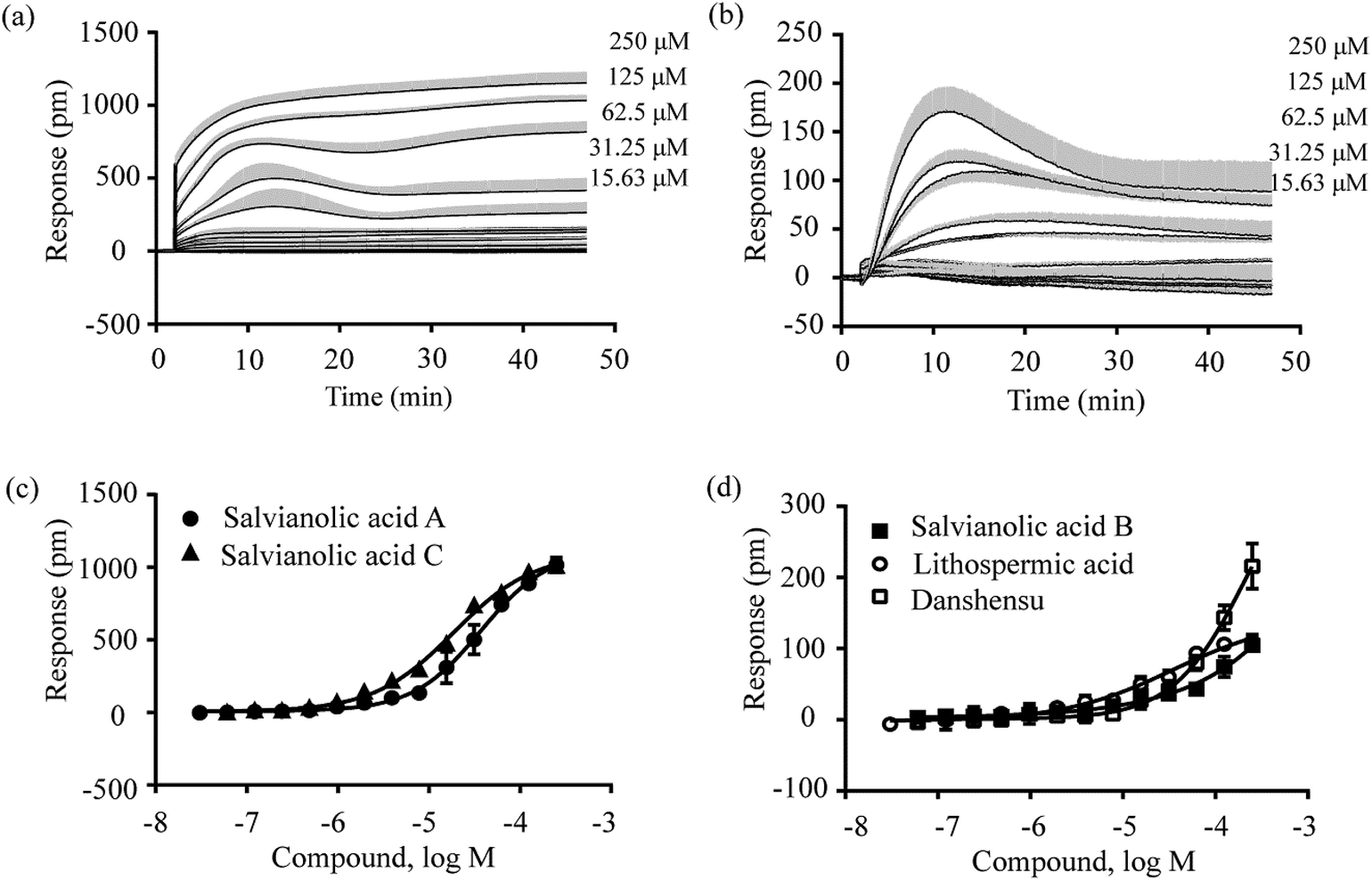
Second, the DMR desensitization assay was performed against the DMR of 1 μM zaprinast. The results showed that these five compounds all dose-dependently desensitized the DMR of zaprinast, leading to apparent IC50 values of 3.48 ± 0.38 μM, 72.74 ± 22.96 μM, 2.70 ± 0.31 μM, 109 ± 23 μM and 30.42 ± 2.25 μM (n = 4 for all) for salvianolic acid A, salvianolic acid B, salvianolic acid C, lithospermic acid and danshensu, respectively (Fig. 5; Table 1). These results suggest that these compounds activate GPR35.
 | ||
| Fig. 5 The DMR amplitudes of 1 μM zaprinast as a function of compound doses in HT-29 cells after pre-stimulation with these compounds for 1 h. (a) Salvianolic acid A and salvianolic acid C; (b) salvianolic acid B, lithospermic acid and danshensu. All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
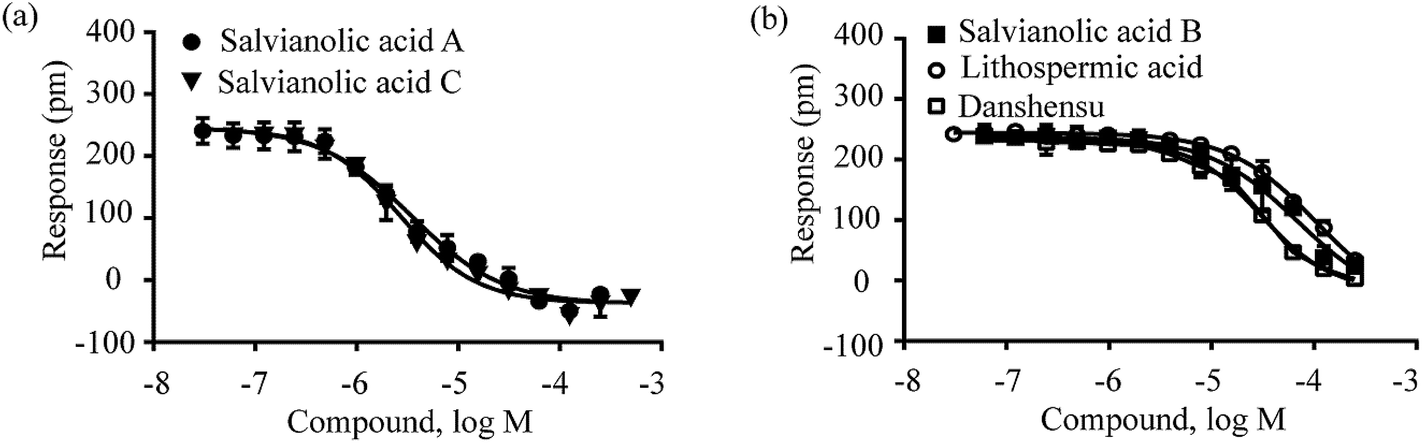
Third, the DMR antagonist assay was performed to confirm the specificity of these compounds for activating GPR35. Known GPR35 antagonists SPB0514229,30 and ML14531,32 were used to pretreat HT-29 cells at different doses, followed by detecting the DMR signal induced by each compound at its respective dose close to the EC80, EC50 or EC20. The results showed that SPB05142 displayed different inhibitory effects on the DMR of different compounds at different doses. When the dose of each compound was assayed at its EC80, SPB05142 dose-dependently and completely blocked the DMR of 125 μM lithospermic acid or 100 μM danshensu, leading to IC50 values of 2.29 ± 0.52 μM and 2.98 ± 1.35 μM (n = 4), respectively (Fig. 6a). However, SPB05142 only partially blocked the DMR of 100 μM salvianolic acid B with an IC50 of 0.71 ± 0.42 μM (n = 4) (Fig. 6a), but had little effect on the DMR of 125 μM salvianolic acid A or 100 μM salvianolic acid C (Fig. 6b). On the other hand, SPB05142 weakly attenuated the DMR of salvianolic acid A and salvianolic acid C at their EC50 values (Fig. 6c), but dose-dependently inhibited their DMR at their respective EC20 values (15 μM and 4 μM, respectively) with IC50 values of 2.52 ± 0.48 μM and 1.57 ± 0.64 μM (n = 4), respectively (Fig. 6d). An almost identical inhibition pattern was observed using ML145 as an antagonist (Fig. S2,† Table 1).
 | ||
| Fig. 6 The DMR amplitudes of compounds as a function of SPB05142 doses in HT-29 cells after being pre-treated with the antagonist for 5 min. (a) Lithospermic acid, salvianolic acid B and danshensu at their EC80 values of 125 μM, 100 μM and 100 μM, respectively; (b) salvianolic acid A and salvianolic acid C at their EC80 values of 125 μM and 100 μM, respectively; (c) salvianolic acid A and salvianolic acid C at their EC50 values of 35 μM and 20 μM, respectively; (d) salvianolic acid A and salvianolic acid C at their EC20 values of 15 μM and 4 μM, respectively. All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
We further examined the activity of salvianolic acid A, salvianolic acid C, salvianolic acid B, lithospermic acid and danshensu using the ERK phosphorylation assay. The activation and signaling of GPR35 induce ERK phosphorylation.33 Fig. 7 shows that salvianolic acid A, salvianolic acid C, salvianolic acid B, lithospermic acid and danshensu lead to ERK phosphorylation (Fig. 7a). As controls, the known GPR35 agonist zaprinast also triggers ERK phosphorylation (Fig. 7b). Moreover, the GPR35 antagonist ML145 attenuated the ERK phosphorylation induced by these compounds (Fig. 7b). These results suggest that salvianolic acid A, salvianolic acid C, salvianolic acid B, lithospermic acid and danshensu resulted in the phosphorylation of ERK via the activation of GPR35.
 | ||
| Fig. 7 ERK phosphorylation assay of active phenolic acids. (a) Western blot of p-ERK1/2 after treatment with salvianolic acid A (SAA), salvianolic acid C (SAC), salvianolic acid B (SAB), lithospermic acid (LA) and danshensu (DSS) at concentrations of 20 μM, 10 μM, 50 μM, 50 μM and 50 μM, respectively. (b) Western blot of p-ERK1/2 after treatment with the known GPR35 agonist zaprinast (Zap) at 1 μM and western blot of p-ERK1/2 after treatment with compounds at the same concentrations as in (a) in the presence of ML145 (25 μM). “Con” represents the blank control. | ||
Together, these results suggest that all five compounds display agonistic activity towards GPR35. Lithospermic acid and danshensu are mostly specific for the activation of GPR35 in HT-29 cells. However, salvianolic acid B may also activate another unknown target, and both salvianolic acid A and salvianolic acid C at high doses may activate multiple targets/pathways, triggering a quite large DMR.
3.3. Deconvolution of possible target(s), besides GPR35, of salvianolic acid A and salvianolic acid C
Both salvianolic acid A and salvianolic acid C appear to display polypharmacology. This was evidenced by the fact that both triggered a larger DMR consisting of contributions from GPR35 as well as other unknown pathway(s) in HT-29 cells. Furthermore, both compounds also triggered a large DMR in A431 cells and attenuated the bradykinin- and histamine-induced DMR signals, probably due to a heterologous desensitization mechanism. The activation of Gq-coupled receptors is known to sequentially activate trimeric G proteins and phospholipase C (PLC), the latter of which then hydrolyzes the membrane lipid phosphatidylinositol bisphosphate, yielding diacylglycerol (DAG) and inositol triphosphate (IP3). IP3 binds to and opens a calcium channel in the endoplasmic reticulum, causing calcium mobilization. On the other hand, danshen is known to cause vasorelaxant actions due to the inhibition of Ca2+ flux.34,35 Therefore, we hypothesized that salvianolic acid A and salvianolic acid C may alter intracellular Ca2+ via an unknown target. To validate this hypothesis, we used thapsigargin and cyclopiazonic acid as the probe molecules. Thapsigargin is a non-competitive inhibitor of sarco/endoplasmic reticulum Ca2+-ATPase (SERCA),36 while cyclopiazonic acid is a specific inhibitor of SERCA.37 Both inhibitors can raise the intracellular calcium concentration by blocking the ability of the cell to pump calcium into the sarcoplasmic and endoplasmic reticula.35The DMR assay of the first step showed that salvianolic acid A and C triggered robust DMR with similar characteristics in A431 cells (Fig. 8a and b), and salvianolic acid C exhibited a slightly higher potency than salvianolic acid A (Fig. 8c). On the other hand, thapsigargin and cyclopiazonic acid triggered comparable DMR in A431 cells, whose characteristics were different from those of salvianolic acid A and C (comparing Fig. 8d and e with Fig. 8a and b). As expected,35 thapsigargin exhibited higher potency than cyclopiazonic acid (Fig. 8f). The IC50 values to trigger DMR were found to be 122.46 ± 11.75 μM, 60.75 ± 3.01 μM, 0.30 ± 0.02 μM and 2.08 ± 0.09 μM (n = 4) for salvianolic acid A, salvianolic acid C, thapsigargin and cyclopiazonic acid, respectively.
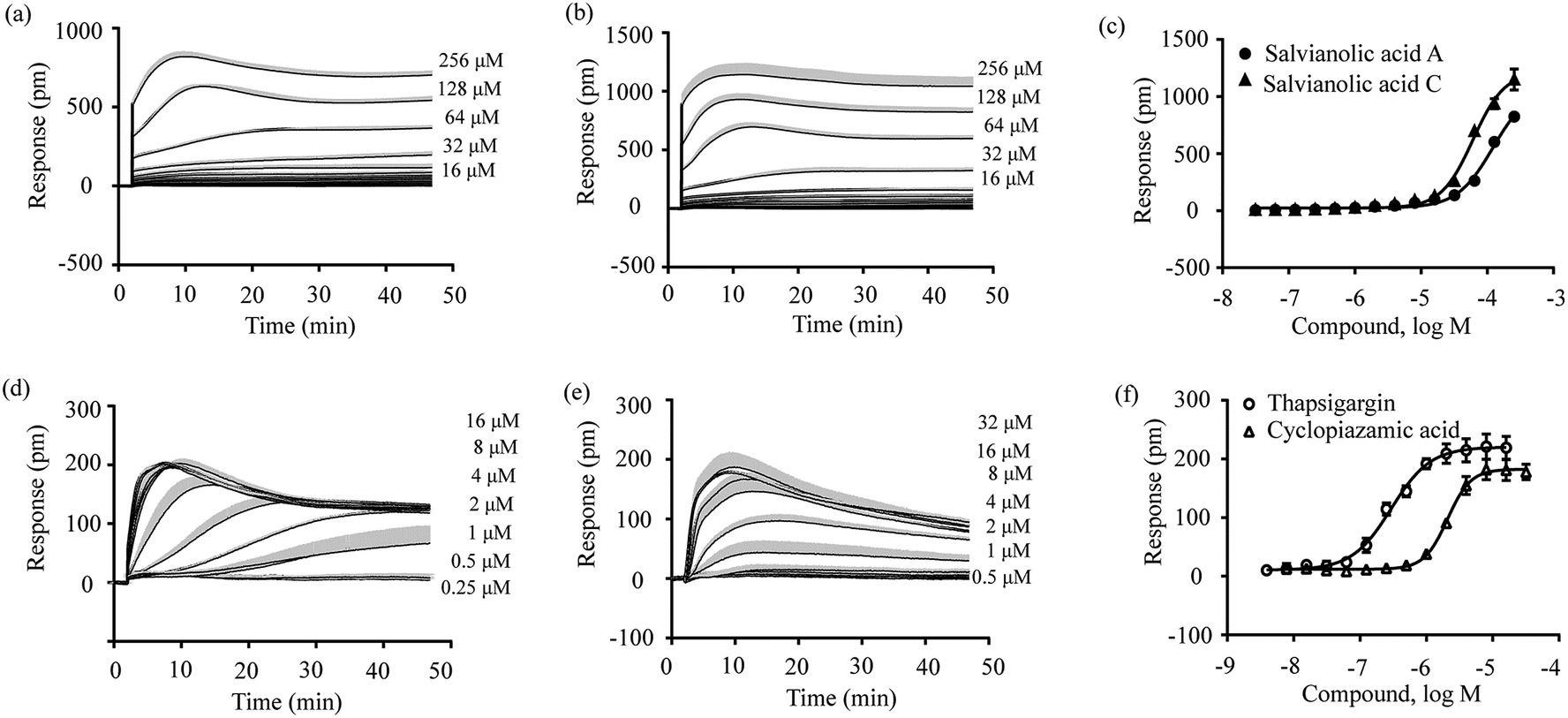 | ||
| Fig. 8 Real time DMR signals of salvianolic acid A (a), salvianolic acid C (b), thapsigargin (d) and cyclopiazonic acid (e) at different doses in A431 cells and their DMR amplitudes as a function of their doses (c and f). All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
The DMR assay of the second step showed that all four compounds caused dose-dependent desensitization of cells responding to the second stimulation with 16 μM cyclopiazolic acid (Fig. 9). The IC50 values to desensitize the cyclopiazolic acid DMR were found to be 71.43 ± 9.53 μM, 26.06 ± 1.70 μM, 0.023 ± 0.002 μM and 5.12 ± 0.50 μM (n = 4 for all) for salvianolic acid A, salvianolic acid C, thapsigargin and cyclopiazonic acid, respectively. These results suggest that salvianolic acid A and salvianolic acid C alter intracellular Ca2+ via an unknown target and share a similar mechanism to those of thapsigargin and cyclopiazonic acid. Further elucidation of the exact target intervened by salvianolic acid A and salvianolic acid C is warranted and is currently under investigation.
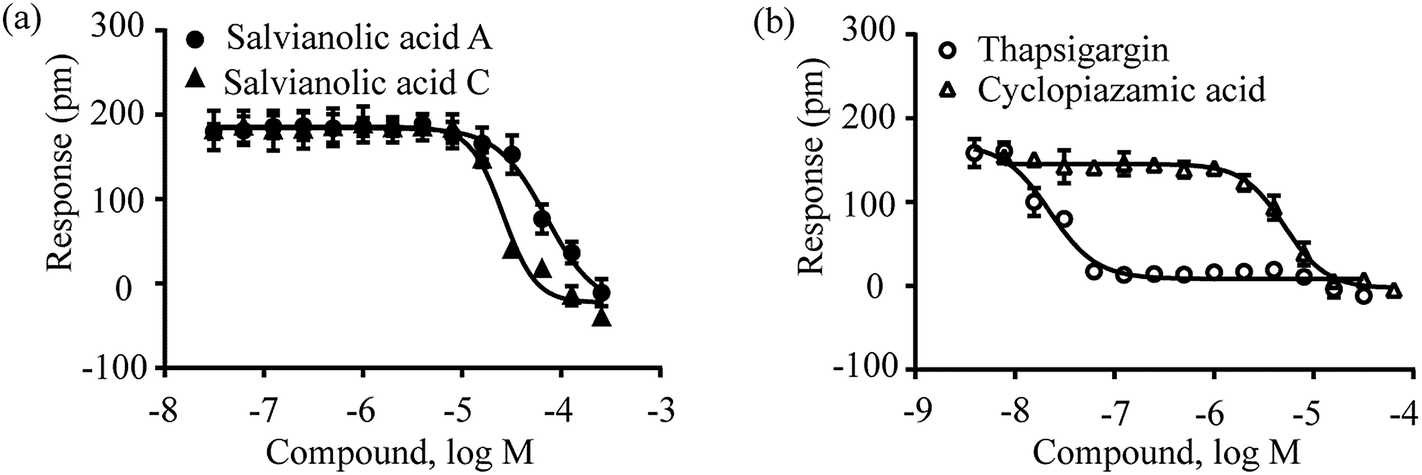 | ||
| Fig. 9 The DMR amplitudes of 16 μM cyclopiazonic acid as a function of compound doses in A431 cells after pre-stimulation with these compounds for 1 h. (a) Salvianolic acid A and salvianolic acid C; (b) thapsigargin and cyclopiazonic acid. All data represents mean ± s.d. from 2 independent measurements, each in duplicate (n = 4). | ||
4. Conclusion
Danshen water extract has clinical benefits for treating cardiovascular and cerebrovascular diseases and contains several phenolic acids. Here, we used a label-free cell phenotypic assay to investigate the possible targets of these phenolic acids from danshen. We found that lithospermic acid, salvianolic acid A, salvianolic acid B, salvianolic acid C and danshensu all displayed agonistic activity against GPR35, but with different potencies. We also found that salvianolic acid A and salvianolic acid C also alter intracellular Ca2+ via a mechanism that is similar to that of two known SERCA inhibitors. The multi-target activity of these phenolic acids may be related to the clinical features of the danshen products. Specifically, GPR35 has been implicated in inflammation, hypertension, coronary artery disease and cancer. Thus, the agonistic activity of multiple phenolic acids in danshen may be useful for elucidating the mechanism of danshen products in the treatment of cardiovascular diseases.Acknowledgements
This work was funded by the Project of National Science Foundation of China (81473436) and the External Cooperation Program of BIC, Chinese Academy of Science (121421KYSB20130013). We also acknowledge the support from the Project of National Key Technology Research and Development Program for The 12th Five-year Plan (2012BAD33B03).References
- D. J. Newman and G. M. Cragg, J. Nat. Prod., 2012, 75, 311–335 CrossRef CAS PubMed.
- M. S. Butler, Nat. Prod. Rep., 2008, 25, 475–516 RSC.
- D. C. Swinney and J. Anthony, Nat. Rev. Drug Discovery, 2011, 10, 507–519 CrossRef CAS PubMed.
- J. Lee and M. Bogyo, Curr. Opin. Chem. Biol., 2013, 17, 118–126 CrossRef CAS PubMed.
- E. Tran and Y. Fang, J. Biomol. Screening, 2008, 13, 975–985 CrossRef CAS PubMed.
- Y. Fang, Expert Opin. Drug Discovery, 2011, 6, 1285–1298 CrossRef CAS PubMed.
- Y. Fang, J. Pharmacol. Toxicol. Methods, 2013, 67, 69–81 CrossRef CAS PubMed.
- Y. Fang, A. M. Ferrie, N. H. Fontaine, J. Mauro and J. Balakrishnan, Biophys. J., 2006, 91, 1925–1940 CrossRef CAS PubMed.
- Y. Fang, Sensors, 2007, 7, 2316–2329 CrossRef CAS PubMed.
- A. M. Ferrie, H. Y. Sun and Y. Fang, Sci. Rep., 2011, 1, 1–8 CrossRef PubMed.
- H. Y. Deng, H. B. Hu and Y. Fang, Sci. Rep., 2012, 2, 1–12 CrossRef PubMed.
- H. Y. Deng, C. M. Wang, M. Su and Y. Fang, Anal. Chem., 2012, 84, 8232–8239 CrossRef CAS PubMed.
- L. M. Zhou, Z. Zuo and M. S. S. Chow, J. Clin. Pharmacol., 2005, 45, 1345–1359 CrossRef CAS PubMed.
- Y. G. Li, L. Song, M. Liu, H. Zhi Bi and Z. T. Wang, J. Chromatogr. A, 2009, 1216, 1941–1953 CrossRef CAS PubMed.
- X. P. Chen, J. J. Guo, J. L. Bao, J. J. Lu and Y. T. Wang, Med. Res. Rev., 2014, 34, 768–794 CrossRef CAS PubMed.
- M.-H. Li, J.-M. Chen, Y. Peng, Q. Wu and P.-G. Xiao, J. Ethnopharmacol., 2008, 120, 419–426 CrossRef PubMed.
- M. Liu, Y. G. Li, F. Zhang, L. Yang, G. X. Chou, Z. T. Wang and Z. B. Hu, J. Sep. Sci., 2007, 30, 2256–2267 CrossRef CAS PubMed.
- R.-W. Jiang, K.-M. Lau, P.-M. Hon, T. C. Mak, K.-S. Woo and K.-P. Fung, Curr. Med. Chem., 2005, 12, 237–246 CrossRef CAS.
- J. H. C. Ho and C. Y. Hong, J. Biomed. Sci., 2011, 18, 1–5 CrossRef.
- J. Huang, Y. Qin, B. Liu, G. Y. Li, L. Ouyang and J. H. Wang, Cell Proliferation, 2013, 46, 595–605 CAS.
- D. G. Kang, H. Oh, H. T. Chung and H. S. Lee, Phytother. Res., 2003, 17, 917–920 CrossRef CAS PubMed.
- B. Jiang, J. Chen, L. Xu, Z. Gao, Y. Deng, Y. Wang, F. Xu, X. Shen and D.-a. Guo, BMC Pharmacol., 2010, 10, 10 CrossRef PubMed.
- B. Sperl, M. H. J. Seifert and T. Berg, Bioorg. Med. Chem. Lett., 2009, 19, 3305–3309 CrossRef CAS PubMed.
- H. Y. Deng, H. B. Hu and Y. Fang, FEBS Lett., 2011, 585, 1957–1962 CrossRef CAS PubMed.
- H. Y. Deng, H. Y. Sun and Y. Fang, J. Pharmacol. Toxicol. Methods, 2013, 68, 323–333 CrossRef CAS PubMed.
- Y. Taniguchi, H. Tonai-Kachi and K. Shinjo, FEBS Lett., 2006, 580, 5003–5008 CrossRef CAS PubMed.
- C. Liebmann, A. Graness, B. Ludwig, A. Adomeit, A. Boehmer, F. D. Boehmer, B. Nurnberg and R. Wetzker, Biochem. J., 1996, 313, 109–118 CAS.
- J. H. Wang, N. Simonavicius, X. S. Wu, G. Swaminath, J. Reagan, H. Tian and L. Ling, J. Biol. Chem., 2006, 281, 22021–22028 CrossRef CAS PubMed.
- P. Zhao, H. Sharir, A. Kapur, A. Cowan, E. B. Geller, M. W. Adler, H. H. Seltzman, P. H. Reggio, S. Heynen-Genel and M. Sauer, Mol. Pharmacol., 2010, 78, 560–568 CrossRef CAS PubMed.
- H. Y. Deng, H. B. Hu, M. Q. He, J. Y. Hu, W. J. Niu, A. M. Ferrie and Y. Fang, J. Med. Chem., 2011, 54, 7385–7396 CrossRef CAS PubMed.
- S. Heynen-Genel, R. Dahl, S. Shi, M. Sauer, S. Hariharan, E. Sergienko, S. Dad, T. Chung, D. Stonich and Y. Su, Probe Reports from the Molecular Libraries Program, 2010, BK5070 Search PubMed.
- H. Deng and Y. Fang, Pharmaceuticals, 2013, 6, 500–509 CrossRef CAS PubMed.
- P. Zhao, H. Sharir, A. Kapur, A. Cowan, E. B. Geller, M. W. Adler, H. H. Seltzman, P. H. Reggio, S. Heynen-Genel, M. Sauer, T. D. Y. Chung, Y. Bai, W. Chen, M. G. Caron, L. S. Barak and M. E. Abood, Mol. Pharmacol., 2010, 78, 560–568 CrossRef CAS PubMed.
- F. F. Y. Lam, J. H. K. Yeung, J. H. Y. Cheung and P. M. Y. Or, J. Cardiovasc. Pharmacol., 2006, 47, 139–145 CrossRef CAS.
- F. F. Y. Lam, J. H. K. Yeung, K. M. Chan and P. M. Y. Or, Vasc. Pharmacol., 2007, 46, 271–277 CrossRef CAS PubMed.
- M. Treiman, C. Caspersen and S. B. Christensen, Trends Pharmacol. Sci., 1998, 19, 131–135 CrossRef CAS.
- K. Moncoq, C. A. Trieber and H. S. Young, J. Biol. Chem., 2007, 282, 9748–9757 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra16102e |
| This journal is © The Royal Society of Chemistry 2015 |