
Lipopeptide biosurfactant from Bacillus clausii BS02 using sunflower oil soapstock: evaluation of high throughput screening methods, production, purification, characterization and its insecticidal activity†
Chinmay Hazra,
Debasree Kundu and
Ambalal Chaudhari*
School of Life Sciences, North Maharashtra University, P.B. No. 80, Jalgaon 425 001, Maharashtra, India. E-mail: ambchasls@gmail.com; ambchasls@yahoo.com; Tel: +91 257 2257425
First published on 27th November 2014
Abstract
A total of 76 bacterial isolates were screened for biosurfactant activity using 16 different high throughput screening methods. Based on phenotypic characteristics, 16S rDNA sequencing and FAME profiling, isolate BS02, identified as Bacillus clausii BS02 produced a biosurfactant (2.6 g l−1) when grown in basal salts mineral medium with sunflower oil soapstock (3% w/v) at 30 °C and 120 rpm after 42 h. The cell free broth showed 82% emulsification activity, critical micelle concentration of 45 mg l−1 and reduced the surface tension from 69.07 mN m−1 to 30 mN m−1. Acid precipitated and methanol extracted crude biosurfactant had 64.3%, 28%, and 4.5% (w/w) of protein, lipid, and carbohydrate, respectively. The crude biosurfactant exhibited stability with oils, fats and non-aqueous phase liquids. Fourier transform infrared (FT-IR) spectroscopy, energy dispersive X-ray spectroscopy (EDX), and nuclear magnetic resonance spectroscopy (NMR) revealed the functional groups and bonds; X-ray diffraction (XRD) and thermogravimetry (TG) showed the surface nature and thermostability of the biosurfactant, respectively. LC-ESI-MS identified the crude biosurfactant as a surfactin lipopeptide. A dose-dependent mortality against C. chinensis and M. hirsutus was observed using the crude biosurfactant, with a LC50 of 50 μg ml−1, indicating the first evidence of its insecticidal activity against pulse beetles and mealybugs.
1. Introduction
Microbial biosurfactants, a heterogeneous group of amphiphilic surface-active compounds produced by bacteria, yeast and filamentous fungi, have received renewed interest in newer production trends and usage patterns due to their low toxicity, high biodegradability, better environmental compatibility, high foaming capability, higher selectivity, specific activity at extreme temperatures, pH, and salinity, and sustainable production from renewable feedstocks.1–5 These properties and the ecological awareness of end-users have led to an upsurge in application of biosurfactants in dairy, food, beverage, cosmetics, detergent, textile, paint, mining, petroleum, paper/pulp, bioremediation, pharmaceuticals, biomedicine and nanotechnology industries.6,7 However, biosurfactants have not yet been commercialized extensively owing to low production yields, high feedstock costs and purification costs and non-availability of non-pathogenic microorganisms.2,3The vegetable oil refineries (∼90) in India manufacture oil from soybean, sunflower, olive, groundnut, rapeseed, safflower, cotton, sesame, coconut, palm, mustard, rice bran, watermelon, etc.8,9 and generate 2–3% (of the total oil production) alkaline (pH 10–12) greasy material (density: 1.7 mg l−1), called soapstock and acidic water (pH 2.6).10 Besides, the volume of wastewater generated is up to 10 l kg−1 of oil (at a rate of about 5% of the volume of refined crude oil) with chemical oxygen demand (COD) value of 2700 mg l−1.10,11 Thus, the use of such low cost by-products (as low as about 10% of the price for refined oil) can (i) stabilize waste disposal by valorisation or minimization, and (ii) produce biosurfactants economically. Several previous literatures revealed that plant and vegetable oils based soapstock are better substrate for rhamnolipid production by P. aeruginosa, accounting for production up to 7.3 g l−1,10 11.6 g l−1,12 and 15.9 g l−1.13 However, efficient biosurfactant production using soapstock by Bacillus spp. has not been reported yet. B. clausii can hold a promise for production of biosurfactants since they are recognized as GRAS (Generally Recognized as Safe) by US FDA and the German Technical Rules for Biological Materials (TRBA).
Hence, the present study attempts to explore high-throughput rational screening methods for isolating an efficient B. clausii isolate BS02, followed by the production of lipopeptide biosurfactants using soapstock from the sunflower oil process as the sole carbon source. Subsequently, the crude biosurfactants was partially purified and characterized. Finally, the insecticidal activity of the crude biosurfactants based on vapor toxicity was evaluated against stored-product grain pest, Callosobruchus chinensis and plant sap-sucking agricultural insect, Maconellicoccus hirsutus.
2. Materials and methods
2.1. Chemicals and reagents
All the reagents, chemicals and media used in this study were purchased from Hi-Media, Mumbai, India until specified. Methanol and chloroform were purchased from Merck, Mumbai, India. All the reagents, chemicals, and media used in this work were of analytical grade. High quality diesel oil was supplied locally by M/s Khubchand Sagarmal (a Hindustan Petroleum franchisee). Crude/raw glycerol from biodiesel co-product stream and sunflower oil soapstock was a generous gift from Jain Irrigation Systems Ltd., Jalgaon and Rising Flora Biotech Pvt. Ltd., Jalgaon, respectively.2.2. Enrichment, isolation and selection of biosurfactant producers
Terrestrial (soil and water) samples were collected from diverse eco-habitats (geographical coordinates: 20.99°N, 75.56°E) and detailed procedure is described in ESI.†2.3. Screening for biosurfactant-producing bacteria using high-throughput rational screening techniques
In order to determine the ability of the isolates to produce biosurfactant, the culture supernatant from each isolate was subjected to preliminary screening methods namely tilted glass slide test, Parafilm M test, drop collapse test, hydrocarbon overlay agar assay, microplate assay, droplet assay, penetration assay, replica plate assay, bacterial adhesion to hydrocarbon (BATH) assay, blood agar hemolysis, CTAB-MB assay, oil spreading assay, emulsification index (EI24) assay, surface tension measurement, contact angle measurements, and atomized oil assay. In all the above mentioned experiments, the cell free culture supernatant of the isolates obtained from Section 2.2 were used following the protocols detailed elsewhere.14–16 Determinations of contact angle are given in ESI.† All the experiments were done in triplicates.2.4. Identification of the selected isolate
Phenotypic characterization, whole-cell fatty acid methyl ester (FAME) profiling, and 16S rDNA sequencing of the selected isolate are described in ESI.†2.5. Fermentative time course of growth and biosurfactant production
A 2% (v/v) actively growing cell suspension of the selected isolate was used to inoculate in a pre-sterile BSM broth containing soapstock (3% w/v) sourced from a sunflower-oil process. Experiments were conducted, in three independent replicates, in a rotary shaker with 500 ml Erlenmeyer flasks containing 100 ml of medium and incubated at 30 °C and 120 rpm. Sample (10 ml) was drawn at appropriate time intervals and monitored for biosurfactant production and biomass growth.2.6. Isolation, partial purification and structural characterization of the biosurfactant
Extracellular crude biosurfactant from a 72 h old culture broth of the isolate was recovered by acidification of the cell free broth and precipitation followed by solvent extraction of the acid precipitate as reported earlier.17 The methanol extracts were subsequently dried and determined gravimetrically.Compositional analysis of the crude sample was determined by ninhydrin test (proteins), anthrone assay (carbohydrate) and saponification test (lipids) according to Patowary et al.18 Surface tension, interfacial tension, critical micelle concentration (CMC) and emulsification was determined as reported previously.19,20 For determining preliminary chemical nature of the biosurfactants, crude sample was spotted on silica gel 60 thin-layer chromatography plates (F254, Merck, Darmstadt Germany). After development in chloroform–methanol–water (65![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :25:4, v/v), lipids were detected with iodine vapors and amino acids/peptides were detected with rhodamine B (250 mg rhodamine B in 100 ml absolute alcohol) and ninhydrin reagent (0.2% ninhydrin solution in acetone). Separation of crude extracts was done using high performance liquid chromatography (HPLC; Agilent 1100 series) equipped with a Zorbax Eclipse column [C18, 5 μm, 4.6 (ID) × 250 mm (L)]. The column was run for 15 min with water–acetonitrile (with 0.1% TCA) (42:58) at a flow rate of 1 ml min−1. Sample injection volume was 50 μl and detection was monitored at 210 nm. UV spectrum of serially diluted (up to 10−3) stock (1 mg ml−1) of the crude biosurfactant (in methanol) was recorded against methanol as a blank in the range of 190–400 nm. Fourier transform infrared spectroscopy (FT-IR) of the dried crude biosurfactant was done by KBr pellet method.17 Energy dispersive spectroscopy (EDS) of the drop-coated grids of vacuum-dried samples (0.5% w/v) was carried out using Bruker attachment (Bruker AXS Ltd., Coventry, UK) with FESEM instrument. The results obtained were reproducible with the accuracy of ±5% error. Powder X-ray diffraction (XRD) patterns were acquired on a Bruker D8 Advance instrument using Cu Kα radiation (λ = 1.5418 Å). A scanning rate of 0.05° s−1 over a wide range of the Bragg angles (θ, 20° ≤ 2θ ≤ 80°) was used. For thermogravimetry (TGA-50, Shimadzu, Tokyo, Japan) and differential scanning calorimetry (DSC-60, Shimadzu, Tokyo, Japan), about 10 mg of dried crude samples were sealed in an Al2O3 crucible with a lid and scanned at a rate of 10 °C min−1 under N2 gas atmosphere (50 ml min−1) over a temperature range of 30–300 °C. The LC-ESI-MS was recorded on a HCT-Ultra ETD II ion trap mass spectrometer (Bruker Daltonics, Germany) coupled to an Agilent 1100 HPLC system. The 1H and 13C NMR spectra of the crude samples (dissolved in CDCl3 at 50 mg ml−1) were collected on a Bruker Avance 300 MHz at room temperature with tetramethylsilane (TMS) as an internal standard.
:25:4, v/v), lipids were detected with iodine vapors and amino acids/peptides were detected with rhodamine B (250 mg rhodamine B in 100 ml absolute alcohol) and ninhydrin reagent (0.2% ninhydrin solution in acetone). Separation of crude extracts was done using high performance liquid chromatography (HPLC; Agilent 1100 series) equipped with a Zorbax Eclipse column [C18, 5 μm, 4.6 (ID) × 250 mm (L)]. The column was run for 15 min with water–acetonitrile (with 0.1% TCA) (42:58) at a flow rate of 1 ml min−1. Sample injection volume was 50 μl and detection was monitored at 210 nm. UV spectrum of serially diluted (up to 10−3) stock (1 mg ml−1) of the crude biosurfactant (in methanol) was recorded against methanol as a blank in the range of 190–400 nm. Fourier transform infrared spectroscopy (FT-IR) of the dried crude biosurfactant was done by KBr pellet method.17 Energy dispersive spectroscopy (EDS) of the drop-coated grids of vacuum-dried samples (0.5% w/v) was carried out using Bruker attachment (Bruker AXS Ltd., Coventry, UK) with FESEM instrument. The results obtained were reproducible with the accuracy of ±5% error. Powder X-ray diffraction (XRD) patterns were acquired on a Bruker D8 Advance instrument using Cu Kα radiation (λ = 1.5418 Å). A scanning rate of 0.05° s−1 over a wide range of the Bragg angles (θ, 20° ≤ 2θ ≤ 80°) was used. For thermogravimetry (TGA-50, Shimadzu, Tokyo, Japan) and differential scanning calorimetry (DSC-60, Shimadzu, Tokyo, Japan), about 10 mg of dried crude samples were sealed in an Al2O3 crucible with a lid and scanned at a rate of 10 °C min−1 under N2 gas atmosphere (50 ml min−1) over a temperature range of 30–300 °C. The LC-ESI-MS was recorded on a HCT-Ultra ETD II ion trap mass spectrometer (Bruker Daltonics, Germany) coupled to an Agilent 1100 HPLC system. The 1H and 13C NMR spectra of the crude samples (dissolved in CDCl3 at 50 mg ml−1) were collected on a Bruker Avance 300 MHz at room temperature with tetramethylsilane (TMS) as an internal standard.
2.7. Insecticidal activities of the crude biosurfactant
The insecticidal activity of the crude biosurfactant was measured by contact toxicity bioassay against adults of stored-product pest insects, the pulse beetle [Callosobruchus chinensis L. (Coleoptera: Bruchidae)] and the mealybug [Maconellicoccus hirsutus (Green) (Hemiptera: Pseudococcidae)]. Rearing of C. chinensis was conducted in plastic boxes (15 × 22 × 10 cm) containing mung beans (Vigna radiata L.) under a photoperiod of 15:9 (L:D) at 25 ± 1 °C and 65 ± 5% relative humidity at Biopesticide Laboratory, School of Life Sciences.21 M. hirsutus collected from an infested Hibiscus rosa-sinensis L. plant in the N.M.U. campus, Jalgaon were reared on the same host plant as per Patil et al.22 For insecticidal activity against adults of C. chinensis, different concentrations of crude biosurfactant (dissolved in methanol) were sprayed on filter papers (Whatman no. 2, 5 cm diameter). For M. Hirsutus, test samples were applied to a small cotton ball attached to the lower end of petiole of healthy leaves of H. rosa-sinensis and air dried. Controls received 500 μl methanol. Each filter paper and leaf was placed in the bottom of a glass Petri plate. Ten test adult insects (1–7 days old) irrespective of sexes were placed on the filter paper or on the leaves in each dish and dish lids replaced. Mortality counts were recorded after every 12 h of exposure at 28 ± 2 °C and relative humidity of 65–70%. Three replicates were run for each treatment and each experiment was repeated thrice on different days to avoid day-to-day variations.
2.8. Statistical analysis
Data are expressed as means ± S.D from at least three independent experiments and were analyzed using Origin 6.1 statistical software (OriginLab Corporation, Northampton, MA, USA). The values of p < 0.05 were considered statistically significant.3. Results and discussion
3.1. Enrichment, isolation and assessment of different high-throughput rational screening techniques for selecting biosurfactant producing isolates
A total of 76 morphologically distinct bacterial isolates, which could grow in BSM supplemented with diesel and crude glycerol (2%, v/v) as sole carbon source were obtained from nine different samples of soil, water, sediment, effluent and sludge (Table S1; ESI†). Of these, 63% of the bacterial isolates were Gram negative. Surprisingly, no filamentous fungi or yeast were found even after three successive enrichments. It may be due to minimal nutritional ingredients [BSM plus diesel and crude glycerol (2%, v/v)] unsupportive for fungal or yeast growth. Among 76 bacterial strains, 54% isolates exhibited biosurfactant activity in the hemolytic assay (Table S2; ESI†). Of these, 13 bacteria showed α-hemolysis while the rest were β-hemolytic. The cell-free culture supernatant of isolate BS01 and BS02 showed distinct and clear hemolysis halos of 2.8 and 2.4 cm, respectively, on blood-agar. Moreover, the diameter of clear zone linearly increased with the concentration of biosurfactant in culture supernatant over a concentration range for the isolate BS01 and BS02 with relatively high R2 values (Fig. S1a; ESI†). Of the 35 nonhemolytic strains, 23 isolates were able to produce biosurfactant both by oil spreading technique and drop collapse method, and 12 other strains showed biosurfactant activity by the oil spreading technique (Table S3; ESI†). On the other hand, out of the 41 positive results obtained with the blood agar method, 16 and 17 gave negative results for biosurfactant production with oil spreading technique and microplate assay, respectively. Thus, the blood agar lysis gave a high percentage of both false positives and negatives. In drop collapse assay, 54 (71%) strains were positive (Table S2; ESI†). Of 22 isolates tested negative by this method (Table S3; ESI†), 8 isolates were also negative for biosurfactant production by oil spreading technique. The other 14 strains were positive for biosurfactant production by the oil spreading technique with diameters of clearing on the oil surface ranging from 0.5 to 0.9 cm, which suggests low levels of biosurfactant concentration (50–63 mg l−1) as per Youssef et al.14 Consistent with this, surface tensions of seven of these strains were high ranging from 55 to 62 mN m−1. Some isolates that were negative for drop collapse activity, but they were all positive for hemolytic and BATH assays. From oil spreading assay, the following general trends could be observed: (i) isolates found with positive drop collapse results were positive for oil spreading assay also; and (ii) the maximum wide clear zone was observed on the oil–water interface (average halo diameter of 4.23 cm to 4.58 cm, respectively) for the isolate BS01 and BS02, respectively and the diameter of the clear zone linearly increased with the concentration of biosurfactant in culture supernatant over a concentration range (Fig. S1b; ESI†). When the degree of oil spreading of 76 isolates was compared to the surface tension of the culture, an inverse linear relationship between the diameter of the clear zone and the surface tension of the culture was obtained (Fig. S1c; ESI†) with a poor correlation coefficient. Wettability alteration measured by contact angle for the isolate BS01 and BS02 were 76° and 75°, respectively, indicating that biosurfactant changed the wettability toward more water-wet, which could be desirable for improving oil recovery at field scale applications. Although the contact angle data correlated well with surface tension measurements, high surface activity of a biosurfactant did not necessarily suggest high emulsifying ability. Out of 76 isolates, 60 and 35 bacteria tested negative for biosurfactant activity with CTAB-MB and blood agar assay, respectively. This integrates the screening procedure and indicates the probable production of anionic surfactant. Table S4 (ESI†) shows the coefficient of correlation between the best five methods (giving maximum no. of positives) that were used to detect biosurfactant production by two different statistical analyses.14 Oil spreading and drop collapse method were strongly correlated with Spearman rank correlation coefficient of rs = 0.91. However, a weak correlation was detected between drop collapse and blood agar lysis method (rs = 0.541) and between oil spreading and blood agar lysis method (rs = 0.453). The statistical test of independence (2 × 2 contingency table) also indicated similar conclusions about the relationships of the five methods in accordance with Youssef et al.14From the data of Tables S2–S4† and Fig. 1, isolate BS02 was selected for subsequent experiments as it showed (i) distinct and clear hemolysis halos (2.4 cm) on blood-agar; (ii) a wide clear zone (average halo diameter of 4.58 cm) on the oil–water interface in OSA; (iii) coalescence over oil covered glass slide in drop collapse test; (iv) highest reduction of surface tension (from 71.14 to 30.06 mN m−1) as compared to other isolates; and (iv) lowest water contact angle (75°).
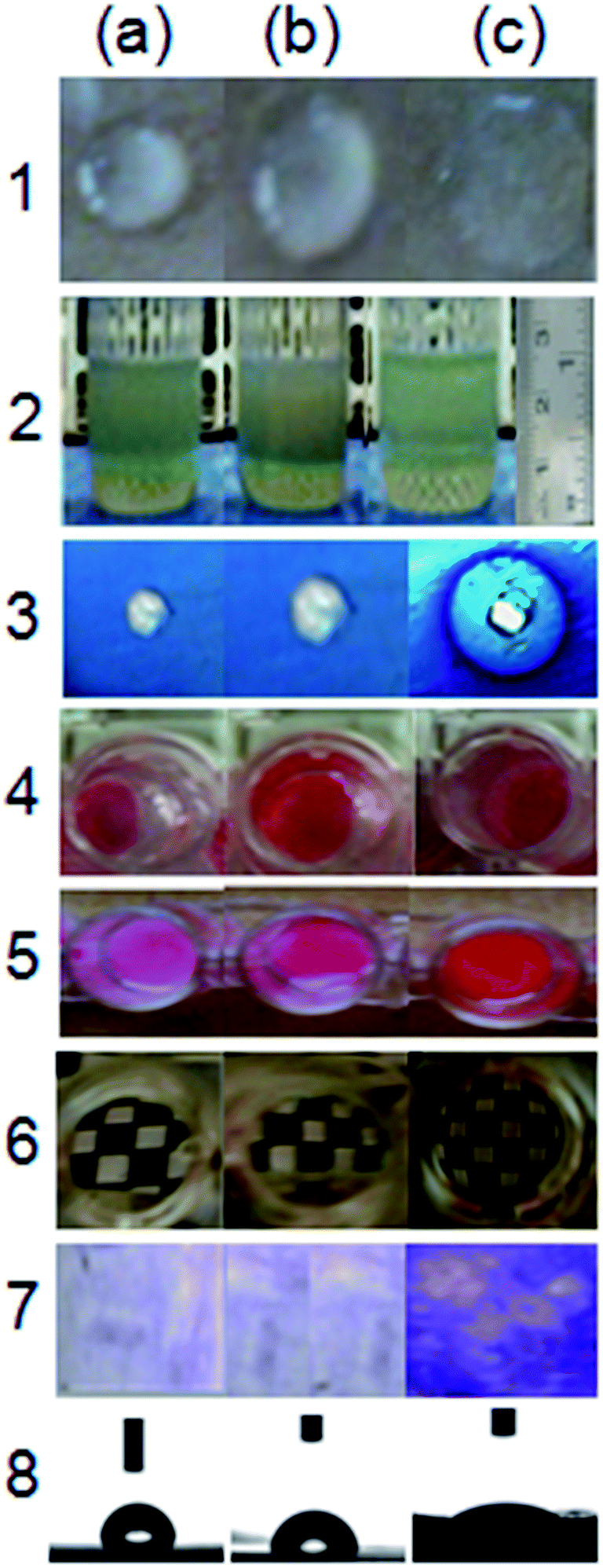 | ||
| Fig. 1 Preliminary screening for detecting biosurfactant activity: (a) Milli-Q water, (b) uninoculated BSM media (with crude glycerol; 2%, v/v), and (c), cell-free culture supernatant from 24 h grown culture of BS02. (1) Parafilm M test; (2) EI24 assay against kerosene; (3) CTAB-MB assay; (4) droplet assay; (5) penetration assay; (6) microplate assay; (7) replica plate assay; (8) water contact angle measurement. All experiments were performed in triplicates. | ||
3.2. Taxonomic affiliation of the biosurfactant producing bacterial strain BS02
Colonies of isolate BS02 were non-pigmented, cream-white, circular, irregular, rough, glistening, erose to lobate, and either slightly raised or low convex. The cells appeared to have translucent, spreading edges with age. The growing cells (on BSM agar supplemented with diesel and/or crude glycerol (2%, v/v) at 30 °C for 48 h) were Gram positive, facultatively aerobic, motile, sporulating, and rod-shaped (0.8–1.0 μm × 2.0–2.5 μm) (Fig. 2A) that grew at pH 6.5–10.0 with an optimal pH of 7.0–8.0, and at 10–41 °C with an optimal temperature of 30 °C. The isolate showed salt tolerance up to 0.2 M NaCl (10% w/v) at pH 9, indicating obligate alkalophile nature. The phenotypic (Table S5; ESI†) and FAME profile (Table S6; ESI†) of isolate BS02 were similar with those of closely related type strains described in the literature, suggesting affiliation to the Bacillus genus.23–25 | ||
| Fig. 2 (A) FESEM photomicrographs of strain BS02 at (a) 7.0k× magnification and (b) 20.0k×. Snap (c) represent Gram staining of BS02 at 100×. (B) Evolutionary relationships of strain BS02 with its relatives inferred using the Neighbor-Joining method. The bootstrap consensus tree inferred from 1000 replicates is taken to represent the evolutionary history of the taxa analyzed. Figures next to the branches represent the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates). The evolutionary distances are in the units of the number of base substitutions per site. Text in the parenthesis is GenBank accession numbers. | ||
For phylogenetic analysis, the almost complete 1422-bp-long 16S rRNA gene was PCR-amplified by using bacterial universal primers, sequenced, and deposited in Gen-Bank under accession number JQ229779. Pairwise sequence analysis and the phylogenetic tree constructed using the neighbour-joining method suggested that strain BS02 is a member of the genus Bacillus and formed a clade with Bacillus clausii KSM K-16 (AB251922) at 100% bootstrap value (Fig. 2B).
3.3. Growth and biosurfactant production profile
The fermentative time-course profile of biosurfactant production by isolate BS02 as a function of time (Fig. 3) depicted that it was detected in the early logarithmic phase of bacterial growth at 24 h. The microbial growth was characterized by a transient lag phase for 18 h followed by an exponential growth phase, which was complete after 54 h of incubation. The biosurfactant production reached to its maximum of 2.6 g l−1 after 42 h of incubation with the YP/X (product yield related to biomass formed) of 1.625 g g−1 and the PV (volumetric productivity) of 0.036 g l−1 h−1. A noticeable decrease in surface tension of the broth culture (from 69.07 mN m−1 to 30 mN m−1) during the exponential growth phase indicated a rationally parallel relationship between biosurfactant production and surface tension reduction (Table S7; ESI†). The reduction in surface tension remained constant until 72 h, when the culture attained the stationary growth phase. Interestingly, a decrease in biosurfactant production was observed after 54 h. It has been reported earlier that in batch fermentation, biosurfactant could be utilized as substrate by the bacteria when the original source of substrate is limited.26 This would explain the sudden drop in the productivity as the isolate approached stationary phase. These findings indicate two aspects: firstly, the biosurfactant biosynthesis occurred predominantly during exponential growth phase, suggesting that (i) the biosurfactant is produced as primary metabolite accompanying the formation of cellular biomass and (ii) the production profile exhibited growth-associated kinetics.26,27 Secondly, biosurfactant production in the presence of soapstock as a sole carbon source may alleviate the growing environmental burden associated with by-product of the vegetable oil processing industry.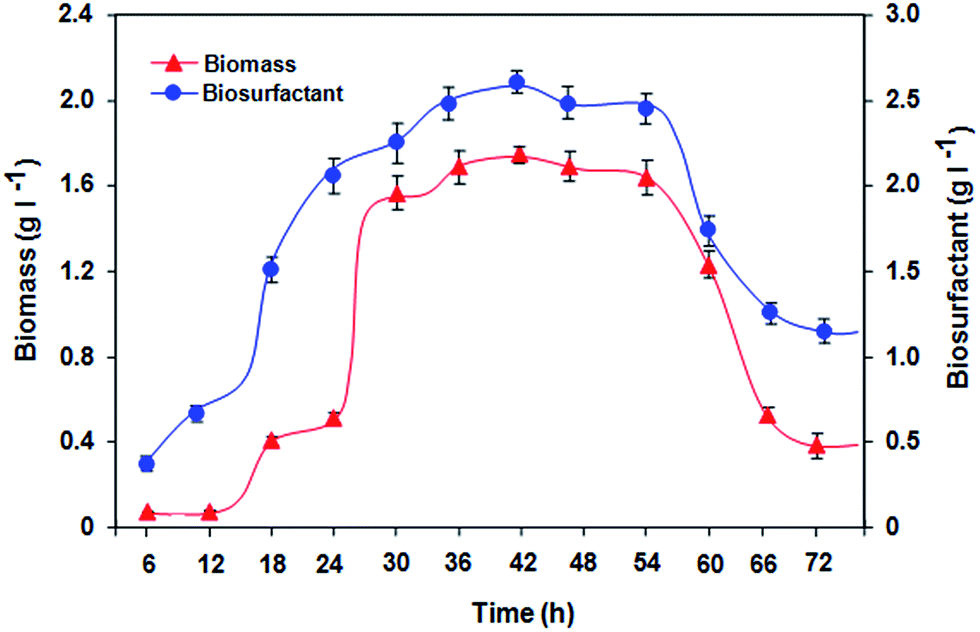 | ||
| Fig. 3 Time course profile of growth of the strain BS02 on BSM medium supplied with sunflower soapstock (3%, w/v) as the sole carbon source accompanied by biosurfactant production. All values are mean ± S.D of triplicates. Small (non-visible) standard deviations are within the symbols. Other process parameters: pH, 8.5; temperature, 30 °C; agitation speed, 120 rpm. | ||
The pH of the culture medium decreased from an initial value of 11.1 ± 0.1 to a maximum of 9.3 ± 0.15 after 48 h of growth, and remained constant thereafter during the subsequent fermentation period. The ability of the isolate to tolerate such alkaline pH during biosurfactant production using soapstock is noteworthy. Earlier literature reports showed that the pH increased from (i) 6.8 to 7.7 for B. subtilis in mineral medium containing purified glycerol,28 (ii) 5.4 to 7.6 for B. subtilis LB5a grown on mineral medium containing cassava wastewater,29 and (iii) 6.9 to 8.0 for B. subtilis LSFM-05 cultivated on minimal medium containing raw glycerol.30
3.4. Partial recovery of the crude biosurfactant
The literature shows different methods for biosurfactant purification, which are dependent on various properties such as acid precipitation, cold acetone extraction, electro-coagulation/flotation, solid phase extraction etc. About 2.5 g l−1 of crude biosurfactants as white solid was isolated from acid precipitated and ethyl acetate extracted 1 l culture of B. clausii BS02.Several strategies have been developed to optimize the production and recovery of purified surfactin using diverse carbon sources, media ingredients, bioreactor design and downstream processes.16 The objective of this work was not to optimize the biosurfactant production but to use the partially purified lipopeptidic compounds as a source of biocontrol agent. Nevertheless, the production yield obtained here is in agreement with the ranges previously reported for surfactin in suboptimal conditions (Table S8 in ESI†). The literature data suggests that B. subtilis ATCC 21332 is a reference strain and it is usually known as a standard surfactin producer in the range of 109.5 to 800.0 mg l−1 depending on the culture medium.31–33 Based on the information summarized in Table S8,† the production of surfactin reported here is lower than previous studies using chemically defined optimized media or using solid-state fermentation. However, it is comparably higher than the earlier literature results obtained with low-cost wastes such as crude oil, biodiesel glycerol, squid pen powder, date molasses, okara and sugarcane bagasse, cashew apple juice, beer wastewater etc. Since one of the motivations of this work was to reduce the costs of producing lipopeptides, the provision of mechanically complex bioreactor designs and labor intensive substrate handling was not considered like other researchers.34 Notably, this is the first report on the use of sunflower oil soapstock as the sole carbon source for surfactin production by B. clausii. This may pave a way for commercial production and bulk applications of surfactin for phytopharmaceutical products since low-cost substrates accounts for 50% of the overall production costs.35
3.5. Characterization of the crude biosurfactant
 | ||
| Fig. 4 (a) RP-HPLC of isolated crude biosurfactant from B. clausii BS02. Fractionation was done as described in text. (b) UV-visible and (c) FTIR spectrum of the crude biosurfactant. | ||
The ESI mass spectra of the purified biosurfactant showed that there were well-resolved group of peaks at m/z values between 1000 and 1100 (Fig. 5a) which may be attributed to the different variants of surfactins, as previously described.30,41 Each surfactin homolog or isoforms can be attributed to the protonated forms, with sodium adducts. The ions of greatest abundances in the sample were those of [M + Na]+ peaks at m/z = 1022, 1030, 1044, 1058 and it correspond to the heptapeptide moiety (L-Glu-L-Leu-D-Leu-L-Val-L-Asp-D-Leu-L-Leu) characteristic of surfactins, linked to a C13, C14 or C15 hydroxy fatty acid chain, respectively.41 The 1H NMR (CDCl3, 300 MHz) spectrum revealed the presence of aldehyde proton at δ = 9.6 ppm; aromatic protons at δ = 7.22–7.68 ppm; a peptide backbone (N–H at 8.0–7.2 ppm) and N-methyl protons at δ = 3.2 ppm (Fig. 5b). The chemical shift at 5.2 ppm detected an ester carbonyl group indicating the presence of a lactone ring in the structure of the biosurfactant. From the 13C NMR (CDCl3, 75.5 MHz) spectrum (Fig. 5c), N-methyl carbon signals was detected at δ = 39.98 ppm, aromatic ring carbon (attached with the carbonyl group) at δ = 154 ppm, and carbonyl signals of the aldehyde group at δ = 185 ppm. A chemical shift at 49.15 ppm indicated a methoxy group attached to a Glu or Asp residue, which is similar with that of the antiadhesin ester and C15-surfactin-O-methyl ester.42 These results showed close resemblance with previous reports of lipopeptide biosurfactant.30
 | ||
| Fig. 5 (a) LC-ESI-MS, (b) 1H NMR spectrum and (c) 13C NMR spectrum (CDCl3 at 25 °C) of crude biosurfactant from B. clausii BS02. | ||
3.6. Insecticidal activities of lipoprotein biosurfactant in insect pests
As shown in Table S10 (ESI†), the treatment of partially recovered biosurfactant led to a dose-dependent mortality against the adults of C. chinensis (L.), reared on mung beans (Vigna radiata L.), producing >50% mortality at 50 μg ml−1 within 48 h and 100% mortality at 100 μg ml−1 within 36 h. Similarly, in case of M. Hirsutus, an exposure period of 36 h was required for 100% mortality at 100 μg ml−1 with LC50 of 50 μg ml−1. The literature survey on microbial bioinsecticides from Gram positive, spore-forming bacterium B. subtilis suggested their (i) mosquito larvicidal and pupicidal activities of cyclic lipopeptides,43 particularly against Anopheles culicifacies, the primary vector of malaria in India; and (ii) larvicidal activities against Mediterranean flour moth (Ephestia kuehniella) (Lepidoptera: Pyralidae),44 Spodoptera littoralis (Lepidoptera: Noctuidae),45 carob moth (Ectomyelois ceratoniae)46 and olive moth (Prays oleae).47 To the best of our knowledge, this is the first report of insecticidal activity of the biosurfactant from B. clausii against C. chinensis (L.) and M. hirsutus adults. Surfactin and its isoforms are cyclic heptalipopeptide class of biosurfactants constituting of both lipophilic and hydrophilic moieties. It is therefore hypothesized that as the insect crawls over the contact surfaces, uptake of biosurfactants may take place through two routes: epicuticular waxes present in the host cuticle, and/or through the intersegmental membranes. This led to leakage or dehydration of the insect cellular components. Nevertheless, the exact mode of action and a field assessment of the insecticidal activity are necessary for the development of formulation of a novel agricultural biopesticide.4. Conclusions
An efficient biosurfactant-producing and crude-oil emulsifying B. clausii BS02, isolated from dairy wastewater is reported using sunflower oil soapstock as the sole carbon and energy source. Putative structural characterization of the crude extract showed it to be mixture of surfactin and its isoforms. The properties of the crude lipopeptide in terms of its CMC, minimum surface tension, interfacial tension, emulsification activity, oil and fat solubilization activity revealed potential applications of the biosurfactant in biotechnological, biomedical and environmental sectors. Being insecticidal to stored-product lepidopteran and plant sap-sucking insect pests, the lipopeptide biosurfactant may be utilized in insect pest management. Further, B. clausii, in contrast to other pathogenic biosurfactant-producers, are classified as GRAS (Generally Recognized as Safe) by US FDA and, therefore, hold potential advantages for their use in industrial scale production and medical applications.Acknowledgements
Chinmay Hazra is grateful to Department of Science and Technology (D.S.T), New Delhi for providing INSPIRE-SRF fellowship. Debasree Kundu is thankful to Council for Scientific and Industrial Research (C.S.I.R), New Delhi for providing CSIR-SRF fellowship. We thank Dr Aniruddha Chatterjee (U.I.C.T., N.M.U., Jalgaon) for helping in instrumental analyses. We express our appreciation to Prof. V. L. Maheshwari, Director, School of Life Sciences, North Maharashtra University, Jalgaon, India and Mr Pankaj Gavit for providing laboratory facilities and useful help in insecticidal assays of C. chinensis. The authors are also thankful to University Grants Commission (U.G.C), New Delhi and Department of Science and Technology (D.S.T), New Delhi for financial support under SAP and FIST program, respectively, to the university.References
- E. J. Gudiña, V. Rangarajan, R. Sen and L. R. Rodrigues, Trends Pharmacol. Sci., 2013, 34, 667–675 CrossRef PubMed.
- P. Bharali, S. P. Singh, S. Gogoi, N. Dutta and B. K. Konwar, RSC Adv., 2014, 4, 38698–38706 RSC.
- P. Saranya, S. Swarnalatha and G. Sekaran, RSC Adv., 2014, 4, 34144–34155 RSC.
- A. Khopade, B. Ren, X.-Y. Liu, K. Mahadik, L. Zhang and C. Kokare, J. Colloid Interface Sci., 2012, 367, 311–318 CrossRef CAS PubMed.
- M. Jadhav, S. Kalme, D. Tamboli and S. Govindwar, J. Basic Microbiol., 2011, 51, 385–396 CrossRef CAS PubMed.
- S. Shekhar, A. Sundaramanickam and T. Balasubramanian, Crit. Rev. Environ. Sci. Technol., 2014 DOI:10.1080/10643389.2014.955631.
- Md. Fakruddin, J. Pet. Environ. Biotechnol., 2012, 3, 124, DOI:10.4172/2157-7463.1000124.
- A. Daverey and K. Pakshirajan, Food Res. Int., 2009, 42, 499–504 CrossRef CAS.
- R. A. Pandey, P. B. Sanyal, N. Chattopadhyay and S. N. Kaul, Resour., Conserv. Recycl., 2003, 37, 101–117 CrossRef.
- M. Benincasa and F. R. Accorsini, Bioresour. Technol., 2008, 99, 3843–3849 CrossRef CAS PubMed.
- I. O. Moraes, M. Benincasa and R. Monte Alegre, Braz. J. Food Technol., 2002, 5, 145–149 Search PubMed.
- M. Benincasa, A. Abalos, I. O. Moraes and A. Manresa, Antonie van Leeuwenhoek, 2004, 85, 1–8 CrossRef CAS PubMed.
- M. Benincasa, J. Contiero, A. Manresa and I. O. Moraes, J. Food Eng., 2002, 54, 283–288 CrossRef.
- N. H. Youssef, K. E. Duncan, D. P. Nagle, K. N. Savage, R. M. Knapp and M. J. McInerney, J. Microbiol. Methods, 2004, 56, 339–347 CrossRef CAS PubMed.
- C.-Y. Chen, S. C. Baker and R. C. Darton, J. Microbiol. Methods, 2007, 70, 503–510 CrossRef CAS PubMed.
- S. K. Satpute, A. G. Banpurkar, P. K. Dhakephalkar, I. M. Banat and B. A. Chopade, Crit. Rev. Biotechnol., 2010, 30, 127–144 CrossRef CAS PubMed.
- P. Das, S. Mukherjee and R. Sen, J. Appl. Microbiol., 2008, 104, 1675–1684 CrossRef CAS PubMed.
- K. Patowary, R. R. Saikia, M. C. Kalita and S. Deka, Ann. Microbiol., 2014 DOI:10.1007/s13213-014-0854-7.
- D. Kundu, C. Hazra, N. Dandi and A. Chaudhari, Biodegradation, 2013, 24, 775–793 CrossRef CAS PubMed.
- C. Hazra, D. Kundu, P. Ghosh, S. Joshi, N. Dandi and A. Chaudhari, J. Chem. Technol. Biotechnol., 2011, 86, 185–198 CrossRef CAS.
- B. K. Salunke, H. M. Kotkar, P. S. Mendki, S. M. Upasani and V. L. Maheshwari, Crop Prot., 2005, 24, 888–893 CrossRef CAS.
- S. V. Patil, C. D. Patil, R. B. Salunkhe, V. L. Maheshwari and B. K. Salunke, Crop Prot., 2011, 30, 1553–1556 CrossRef.
- Y. Nogi, H. Takami and K. Horikoshi, Int. J. Syst. Evol. Microbiol., 2005, 55, 2309–2315 CrossRef CAS PubMed.
- N. K. Asha Devi, K. Balakrishnan, R. Gopal and S. Padmavathy, Curr. Sci., 2008, 95, 627–636 Search PubMed.
- G. Mouloud, H. Daoud, J. Bassem, I. L. Atef and B. Hani, Appl. Biochem. Biotechnol., 2013, 171, 2186–2200 CrossRef CAS PubMed.
- R. Chandankere, J. Yao, M. M. F. Choi, K. Masakorala and Y. Chan, Biochem. Eng. J., 2013, 74, 46–53 CrossRef CAS.
- C. D. Cunha, M. Rosario, A. S. Rosado and S. G. F. Leite, Process Biochem., 2004, 39, 2277–2282 CrossRef CAS.
- F. A. S. L. Reis, E. F. C. Servulo and F. P. de Franca, Appl. Biochem. Biotechnol., 2004, 115, 899–912 CrossRef.
- F. F. C. Barros, A. N. Ponezi and G. M. Pastore, J. Ind. Microbiol. Biotechnol., 2008, 35, 1071–1078 CrossRef CAS PubMed.
- A. F. de Faria, D. S. Teodoro-Martinez, G. N. O. Barbosa, B. G. Vaz, I. S. Silva, J. S. Garcia, M. R. Totola, M. N. Eberlin, M. Grossman, O. L. Alves and L. R. Durrant, Process Biochem., 2011, 46, 1951–1957 CrossRef.
- D. G. Cooper, C. R. Macdonald, S. J. B. Duff and N. Kosaric, Appl. Environ. Microbiol., 1981, 42, 408–412 CAS.
- F.-C. Hsieh, M.-C. Li, T.-C. Lin and S.-S. Kao, Curr. Microbiol., 2004, 49, 186–191 CrossRef CAS PubMed.
- M. Sousa, V. M. M. Melo, S. Rodrigues, H. B. Sant'ana and L. R. B. Gonçalves, Bioprocess Biosyst. Eng., 2012, 35, 897–906 CrossRef CAS PubMed.
- C. T. Slivinski, E. Mallmann, J. Magali de Araújo, D. A. Mitchell and N. Krieger, Process Biochem., 2012, 47, 1848–1855 CrossRef CAS.
- C. Mulligan and B. Gibbs, in Biosurfactants: production, properties, applications, ed. N. Kosaric, Marcel Dekker, NY, 1993, pp. 329–371 Search PubMed.
- K. Das and A. K. Mukherjee, Appl. Microbiol. Biotechnol., 2005, 69, 192–195 CrossRef CAS PubMed.
- S. Joshi, C. Bharucha and A. J. Desai, Bioresour. Technol., 2008, 99, 4603–4608 CrossRef CAS PubMed.
- R. P. Singh, M. K. Shukla, A. Mishra, P. Kumari, C. R. K. Reddy and B. Jha, Carbohydr. Polym., 2011, 84, 1019–1026 CrossRef CAS.
- W. Ismail, I. S. Al-Rowaihi, A. A. Al-Humam, R. Y. Hamza, A. M. El Nayal and M. Bououdina, Int. Biodeterior. Biodegrad., 2013, 84, 168–178 CrossRef CAS.
- R. Chandankere, J. Yao, M. Cai, K. Masakorala, A. K. Jain and M. M. F. Choi, Fuel, 2014, 122, 140–148 CrossRef CAS.
- J. F. B. Pereira, E. J. Gudiña, R. Costa, R. Vitorino, J. A. Teixeira, J. A. P. Coutinho and L. R. Rodrigues, Fuel, 2013, 111, 259–268 CrossRef CAS.
- X.-Y. Liu, S.-Z. Yang and B.-Z. Mu, Process Biochem., 2009, 44, 1144–1151 CrossRef CAS.
- I. Geetha and A. M. Manonmani, Lett. Appl. Microbiol., 2010, 51, 406–412 CrossRef CAS PubMed.
- D. Ghribi, M. Elleuch, L. M. Abdelkefi and S. Ellouze-Chaabouni, J. Stored Prod. Res., 2012, 48, 68–72 CrossRef CAS.
- D. Ghribi, L. Abdelkefi, H. Boukadi, M. Elleuch, S. Ellouze-Chaabouni and S. Tounsi, J. Invertebr. Pathol., 2012, 109, 183–186 CrossRef CAS PubMed.
- I. Mnif, M. Elleuch, S. E. Chaabouni and D. Ghribi, Crop Prot., 2013, 50, 66–72 CrossRef CAS.
- D. Ghribi, I. Mnif, H. Boukadi, K. Radhouan and S. Chaabouni-Ellouze, Afr. J. Microbiol. Res., 2011, 5, 4927–4936 Search PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra13261k |
| This journal is © The Royal Society of Chemistry 2015 |