
Electrocatalytic cycle of P450 cytochromes: the protective and stimulating roles of antioxidants
Abstract
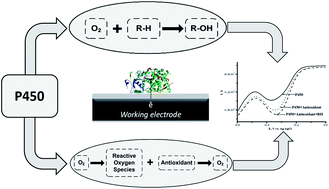
The electrochemical activity of cytochrome P450 (CYP) 3A4, 2C9 and 2D6 on a screen-printed graphite electrode nanostructured with gold nanoparticles and stabilized with didodecyldimethylammonium bromide was examined. The analysis of the CYP catalytic activity was carried out using a variety of electrochemical techniques, such as cyclic voltammetry, square wave voltammetry and amperometry. A sensitive electrochemical CYP-sensor system was proposed as a reliable candidate for investigating the influence of medicinal preparations – ethoxidol (2-ethyl-6-methyl-3-hydroxypyridine malate), mexidol (2-ethyl-6-methyl-3-hydroxypyridine succinate), cytochrome c and L-carnitine – on the CYP redox behavior and electrocatalytic activity. In the presence of antioxidants, the enhancement of the electrochemical signal of CYP was registered. It was shown that ethoxidol, in the concentration range of 90–600 μM, stimulates the electrochemical reduction of P450 cytochromes, with the maximum stimulating effects for CYP3A4, CYP2C9 and CYP2D6 being estimated as 181 ± 15%, 240 ± 7% and 110 ± 4% (at 540 μM ethoxidol), respectively.
 Please wait while we load your content...
Please wait while we load your content...

