Effects of 3D nanocomposite bioceramic scaffolds on the immune response
Mónica
Cicuéndez
ab,
Pilar
Portolés
cd,
María
Montes-Casado
d,
Isabel
Izquierdo-Barba
ab,
María
Vallet-Regí
*ab and
María Teresa
Portolés
*e
aDepartamento de Química Inorgánica y Bioinorgánica, Facultad de Farmacia, Universidad Complutense de Madrid, Plaza Ramón y Cajal s/n, 28040 Madrid, Spain. E-mail: vallet@ucm.es
bNetworking Research Center on Bioengineering, Biomaterials and Nanomedicine (CIBER-BBN), Madrid, Spain
cConsejo Superior de Investigaciones Científicas, CSIC, Spain
dCentro Nacional de Microbiología, Instituto de Salud Carlos III, Majadahonda, 28220-Madrid, Spain
eDepartamento de Bioquímica and Biología Molecular I, Facultad de Ciencias Químicas, Universidad Complutense de Madrid, Ciudad Universitaria s/n, 28040-Madrid, Spain. E-mail: portoles@quim.ucm.es
First published on 31st March 2014
Abstract
The interaction of new nanocomposite mesoporous glass/hydroxyapatite (MGHA) scaffolds with immune cells involved in both innate and acquired immunity has been studied in vitro as an essential aspect of their biocompatibility assessment. Since the immune response can be affected by the degradation products of bioresorbable scaffolds and scaffold surface changes, both processes have been evaluated. No alterations in proliferation and viability of RAW-264.7 macrophage-like cells were detected after culture on MGHA scaffolds which did not induce cell apoptosis. However, a slight cell size decrease and an intracellular calcium content increase were observed after contact of this cell line with MGHA scaffolds or their extracts. Although no changes in the percentages of RAW cells with low and high contents of reactive oxygen species (ROS) are observed by the treatment with 7 day extracts, this study has revealed modifications of these percentages after direct contact with scaffolds and by the treatment with 24 h extracts, related to the high reactivity/bioactivity of this MGHA nanocomposite at initial times. Furthermore, when normal fresh murine spleen cells were used as an experimental model closer to physiological conditions, no significant alterations in the activation of different immune cell subpopulations were detected in the presence of 24 h MGHA extract. MGHA scaffolds did not affect either the spontaneous apoptosis or intracellular cytokine expression (IL-2, IL-10, IFN-γ, and TNF-α) after 24 h treatment. The results obtained in the present study with murine immune cell subpopulations (macrophages, lymphocytes B, lymphocytes T and natural killer cells) support the biocompatibility of the MGHA material and suggest an adequate host tissue response to their scaffolds upon their implantation.
1. Introduction
Tissue engineering, due to its highly multidisciplinary nature, needs the combination of clinical medicine, mechanical engineering, materials science, genetics, and related disciplines from both engineering and life sciences.1 This field relies on the use of porous 3D scaffolds to provide an appropriate environment for the regeneration of tissues and organs.2 These scaffolds essentially act as templates for tissue formation and are seeded with cells and occasionally growth factors. The combination of cells, biomolecules and scaffolds constitutes the tissue engineering triad.3The composition, macroporous architecture, microporosity, surface topography and chemistry, as well as the mechanical properties of scaffolds used for tissue engineering, play a crucial role in the success rates of implants.3 Fundamental aspects in the manufacturing process of 3D scaffolds are the starting material and the methodology which must be used for configuring the 3D structure. Different biomaterials are employed in the fabrication of scaffolds for tissue engineering, with specific advantages and disadvantages. Recently, the use of composite scaffolds allowed improvement of their properties due to the synergy of the features of their components. For this reason, composite scaffolds are widely used for different applications.4–8 In this sense, our research group has recently synthesized a novel nanocomposite material formed by nanocrystalline apatite uniformly embedded into a mesostructured SiO2–CaO–P2O5 glass wall (MGHA material).9 The response of L929 fibroblasts and Saos-2 osteoblasts to this powdered nanocomposite has been previously studied, demonstrating a significantly improved biocompatibility in comparison with conventional mesoporous glasses.9,10 Furthermore, recently 3D scaffolds based on this nanocomposite material have exhibited good preosteoblast cell adhesion, colonization, proliferation and differentiation, suggesting their great potential for application in bone tissue engineering.11
The first characteristic of any scaffold for tissue engineering is that it must be biocompatible, allowing cell adhesion, proliferation and migration through it.12,13 However, the ultimate functional success or failure of the scaffolds after implantation depends invariably on the host tissue response and therefore, the scaffold must not induce an immune reaction which might reduce healing or produce rejection by the body. Thus, the in vitro evaluation of biomaterial effects on the immune system is an essential aspect of biocompatibility assessment.14 The immune response comprises both innate and adaptive defence mechanisms which activate different cell populations co-ordinately. The innate response involves the action of neutrophils, monocytes and macrophages which carry out phagocytosis and produce reactive oxygen species, antimicrobial peptides, and inflammatory mediators.15 Natural killer (NK) cells also play a role in the innate response by directly killing virus-infected or transformed cells without prior priming by secretion of cytotoxic molecules and cytokines.16 The adaptive response is mediated by antigen-specific lymphocytes (T and B cells) which produce inflammatory cytokines and antibodies.15 Largely, the interaction of these host immune cells with biomaterials has negative implications for tissue remodelling. For decades, the interaction of macrophages with a newly deposited extracellular matrix, accompanied by an angiogenic response, has been interpreted as a precursor of granulation tissue formation, the classic foreign body reaction, with negative implications for outcomes.17 However, it has been recently shown that macrophages may be beneficial for biomaterial integration, thereby improving implant performance due to a remarkable functional plasticity. Thus, macrophages can play both positive and negative roles in disease processes and tissue remodeling, presenting a phenotypic spectrum based on three fundamental homeostatic activities, including host defence, wound healing, and immune regulation.18 There are two extremes of possible macrophage phenotypes based on in vitro treatment with particular stimuli: proinflammatory (M1) and reparative (M2),17–20 whose balance determines the host response to biomaterials, with participation of diverse specific cytokines.
In this sense, the design of biomaterials capable of modulating immune cell function has been suggested in order to obtain appropriate host responses at the implantation site.16–22
Given the excellent biocompatibility and bioactivity shown by the MGHA nanocomposite and taking into account its potential application in bone tissue engineering,9,11 the present study is focused on the in vitro interaction of 3D MGHA scaffolds with immune cells involved in both innate and acquired immunity. Furthermore, since the immune response can be additionally affected by the degradation products of bioresorbable scaffolds, the possible cytotoxic effects caused by MGHA extracts obtained at different times have been also studied.
2. Materials and methods
2.1. Preparation of MGHA 3D scaffolds
MGHA 3D scaffolds based on a highly mesostructured nanocomposite material were prepared by rapid prototyping (RP) using methylcellulose (MC), as previously reported.11 Once fabricated, these MGHA scaffolds were sterilized under UV light for 30 min and then submerged in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with penicillin (800 μg ml−1) and streptomycin (800 μg ml−1), under a CO2 (5%) atmosphere at 37 °C for 24 h for stabilization before cell culture.2.2. Characterization of MGHA 3D scaffolds
X-ray diffraction (XRD) experiments were performed on a Philips X'Pert diffractometer (Eindhoven, The Netherlands), equipped with Cu Kα (40 kV, 20 mA). Transmission electron microscopy (TEM) was performed on a JEOL 3010 electron microscope (Jeol Ltd., Japan) operating at 300 kV (Cs; 0.6 mm, resolution 1.7 Å). All TEM images were recorded employing a CCD camera (MultiScan model 794, Gatan Inc., UK) under low-dose conditions. Fourier transform (FT) patterns were extracted from the images of thin crystal regions using a Digital Micrograph (Gatan Inc., UK). Scanning electron microscopy (SEM) was performed using a field emission JEOL JSM-6335F microscope (Tokyo, Japan) at an acceleration voltage of 10 kV. The textural properties of samples were determined by N2 adsorption porosimetry using a Micromeritics ASAP2020 analyzer (Norcross, USA). The surface area was determined using the multipoint Brunauer–Emmett–Teller method included in the software. The porosity of 3D scaffolds was measured by mercury intrusion porosimetry using a MicromeriticsAutopore IV 9500 device (Micromeritics Instrument Corporation, Norcross, GA, USA). Elemental analyses (C, H, and N) were carried out on a LECO CHNS-932 microanalyzer (Saint Joseph, Michigan USA). Fourier transform Infrared (FTIR) spectroscopy was performed using a Thermo Nicolet Nexus spectrometer (Thermo Scientific, USA) from 4000 to 400 cm−1, using the KBr pellet method and operating in transmittance mode.2.3. In vitro degradation of MGHA 3D scaffolds and preparation of MGHA extracts
MGHA scaffolds of ca. 23–25 mg were immersed at different time periods for 30 days in a total volume of 100 ml of DMEM supplemented with 10% fetal bovine serum (FBS) and antibiotics (100 U ml−1 penicillin, 100 mg ml−1 streptomycin), in an Ecotron at 37 °C and 150 rpm. The supernatants obtained after 24 h and 7 days, named E 24 h and E 7 d, were selected for immune response studies. These extracts were filtered and added to RAW-264.7 macrophage-like cells to evaluate any potential cytotoxic effect. Determination of Si(IV), Ca(II) and P(V) variations in these extracts was carried out at different time intervals by inductively coupled plasma/optical emission spectrometry (ICP/OES) using a Perkin Elmer OPTIMA 3300 DV device. The scaffold surface was evaluated by scanning electron microcopy (SEM) after different times.2.4. Culture and treatment of RAW-264.7 macrophage-like cells
Murine RAW-264.7 macrophage-like cells were seeded at a density of 105 cells per ml in a culture medium supplemented with 10% FBS, 1 mM L-glutamine, penicillin, and streptomycin, under a 5% CO2 atmosphere, at 37 °C for 24 h. 24 h and 7 day extracts have been also added to RAW macrophage-like cells to know the effects of the released products during the degradation process. Then, the attached cells were harvested with PBS/1 mM EDTA at 4 °C and counted in a Neubauer hemocytometer. Cell suspensions were centrifuged at 310 × g for 10 min and resuspended in a fresh medium for the analysis of different parameters by flow cytometry as described below (Sections 2.5–2.8). Controls without treatment were carried out in parallel.2.5. Cell cycle analysis and apoptosis detection
After detachment of RAW-264.7 macrophage-like cells, cell suspensions were incubated with Hoechst 33258 (5 μg ml−1, ethanol 30%, and BSA 1% in PBS), used as a nucleic acid dye, for 30 min at room temperature in darkness. The fluorescence of Hoechst was excited at 350 nm and the emitted fluorescence was measured at 450 nm using a LSR Becton Dickinson flow cytometer. The cell percentage in each cycle phase (G0/G1, S and G2/M) was calculated with the CellQuest Program of Becton Dickinson and the SubG1 fraction (cells with fragmented DNA) was used as an indicator of apoptosis.2.6. Cell size and complexity
After detachment of RAW-264.7 macrophage-like cells, forward angle (FSC) and side angle (SSC) scatters were evaluated as indicators of cell size and complexity respectively using a FACScalibur Becton Dickinson flow cytometer.2.7. Intracellular reactive oxygen species (ROS) content and cell viability
After detachment of RAW-264.7 macrophage-like cells, cell suspensions were incubated with 100 μM 2′,7′-dichlorofluorescein diacetate (DCFH/DA) at 37 °C for 30 min. DCF fluorescence was excited at 488 nm and measured with a 530/30 nm band pass filter using a FACScalibur Becton Dickinson flow cytometer. Cell viability was determined by addition of propidium iodide (PI; 0.005% in PBS).2.8. Intracellular calcium content
After detachment of RAW-264.7 macrophage-like cells, cell suspensions were incubated with the probe Indo-1 AM at a concentration of 10 μM for 30 min at room temperature, darkness, and shaking. The fluorescence of Indo-1 was excited at 325 nm and the emitted fluorescence was measured with 380 nm long pass (FL1) and 424/44 nm band pass (FL2) filters using a LSR Becton Dickinson flow cytometer. After all the measurements, 10 μM A-23187 ionophore (Enzo Life Sciences) was added in order to prove the sensitivity of the assay.2.9. Morphological studies by confocal microscopy
For confocal microscopy studies, RAW-264.7 macrophage-like cells were cultured for 24 h on MGHA disks (10 mm diameter and 4 mm high) prepared by uniaxial and isostatic pressure of powder. After fixation with 3.7% paraformaldehyde in PBS for 10 min, samples were washed with PBS and permeabilized with 0.1% Triton X-100 for 3 to 5 min. The samples were then washed with PBS and preincubated with PBS containing 1% BSA for 20 to 30 min. Then cells were incubated for 20 min with FITC phalloidin which stains F-actin filaments. Samples were then washed with PBS and the cell nuclei were stained with DAPI (4′-6-diamidino-2′-phenylindole, 3 μM in PBS). After staining and washing with PBS, cells were examined using a LEICA SP2 Confocal Laser Scanning Microscope. The fluorescence of FITC was excited at 488 nm and the emitted fluorescence was measured at 491–586 nm. The fluorescence of DAPI was excited at 405 nm and measured at 420–480 nm.2.10. Mice
Mice from the C57BL/6 strain were bred under specific pathogen-free conditions in the animal care facility of the Centro Nacional de Microbiología (Instituto de Salud Carlos III, Majadahonda, Madrid, Spain), from stock purchased from Charles River. Sex-matched, 10–12 week old mice were used throughout the experiments. All procedures were approved by Institutional Animal Care and Use Committees.2.11. Culture and activation of murine spleen cells
Cell suspensions from C57Bl/6 spleen in Click's culture medium supplemented with 10% FBS were obtained, and the erythrocytes were lysed by hypotonic shock. Cell suspensions were filtered through a 30 micron mesh and washed. In vitro primary stimulation was performed by culturing 5 × 105 cells in 200 μl of Click's medium supplemented with 10% FBS and 10 mM HEPES, in the presence of Y-CD3-1 (anti-CD3 monoclonal antibody, 5 μg ml−1)23 or lipopolysaccharide (LPS, E. coli 055:B5, 25 μg ml−1) as a stimulus and in the presence or absence of filtered MGHA extracts. After 3 day culture at 37 °C and 5% CO2, viability measured as mitochondrial activity was assessed by a colorimetric assay using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) as a substrate for mitochondrial dehydrogenases in living cells as previously described by Mosmann.242.12. Detection of murine lymphocyte spontaneous apoptosis
To assess the effect of MGHA scaffolds on murine lymphocyte spontaneous apoptosis, spleen cells were prepared as above but cells were cultured in flat-bottom P-48 culture plates containing previously stabilized scaffolds. 5 × 106 cells were seeded onto the scaffold in a final volume of 0.5 ml culture medium. After 24 h culture at 37 °C and 5% CO2, cells were collected and spontaneous apoptosis was determined by flow cytometry in cells stained with Annexin V-FITC and propidium iodide using the Human Annexin V-FITC kit (eBioscience) according to the manufacturer's instructions and analyzed using a FacsCanto (B-D Biosciences) flow cytometer with DIVA software.2.13. Analysis of intracellular cytokine expression
To analyze the effect of MGHA scaffolds on cytokine expression, suspensions of murine spleen cells were prepared as above. 106 cells were seeded onto scaffolds in P-48 culture plates and cultured for 3 days in the presence of Y-CD3-1 (anti-CD3 antibody, 5 μg ml−1) or LPS (50 μg ml−1) or culture medium, in 1 ml final volume. After this time, cells were collected and processed for intracellular cytokine detection by fluorescence-activated cell sorting (FACS). Intracellular cytokine staining was performed essentially as described by Rojo et al.25 Cells were washed and re-suspended at 2 × 106 ml−1 in the culture medium and stimulated with 20 ng ml−1 phorbol dibutyrate (PDB; Sigma-Aldrich) plus 1 μM ionomycin (Calbiochem) for 1.5 h. Brefeldin A (10 μg ml−1; Sigma-Aldrich) was added, and the cells were incubated for further 3.5 h, washed and surface labeled with desired antibodies by incubation for 30 min on ice with fluorocrome-coupled antibodies diluted in cold staining buffer (PBS containing 2% FCS and 0.1% sodium azide). Then, the cells were washed two times and fixed for 5 min at room temperature with 4% paraformaldehyde in PBS. After stopping the fixation with ice-cold 0.1% BSA in PBS/0.1% azide, the cells were washed with 0.1% saponin (Sigma-Aldrich) in PBS/0.1% BSA/10 mM HEPES/azide (PBS/saponin) and blocked at 4 °C for 30 min with 5% non-fat milk in PBS/saponin. Then, the cells were stained with PE-, Alexa Fluor 647-, DyLight 649 (Pierce)- or FITC-coupled antibodies in 5% non-fat milk in PBS/saponin for 30 min in the cold. After washing three times with PBS/saponin, the cells were analyzed using a FACSCanto (B-D Biosciences) flow cytometer. The antibodies for cytokine intracellular detection were from eBioscience: PE-coupled anti-IL-2 (JES6-5H4), and anti-IL-10 (JES5-16E3); FITC-coupled anti-IFNγ (XMG 1.2) and anti-TNFα. As negative controls, PE- or FITC-conjugated isotype control mAb were used.2.14. Statistics
Data are expressed as mean ± standard deviation of a representative of three experiments carried out in triplicate. Statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS) version 19 software. Statistical comparisons were made by analysis of variance (ANOVA). The Scheffé test was used for posthoc evaluations of differences among groups. In all of the statistical evaluations, p < 0.05 was considered as statistically significant.3. Results and discussion
3.1. MGHA 3D scaffolds
The new mesoporous glass/hydroxyapatite nanocomposite MGHA has been used as the starting material for manufacturing the 3D scaffolds by the RP technique, as reported previously.11 Previous results demonstrate its high biocompatibility and bioactivity9,11 and drug loading capability,26 characteristics which make MGHA a potential biomaterial for bone tissue engineering. In the present study, hierarchical 3D MGHA scaffolds and their degradation products (extracts) have been prepared to evaluate their effects on immune cells, involving macrophages and lymphocytes.As it can be observed in Fig. 1, MGHA 3D scaffolds present a hierarchical porous macro–meso architecture which improves cell adhesion, proliferation differentiation and colonization.11
 | ||
| Fig. 1 SEM and TEM studies of the macro and mesostructures of MGHA 3D scaffolds. (A) Cross-section of a 3D scaffold showing the hierarchical pore structure of MGHA scaffolds. The small arrow shows the interconnections of large channels of 400 μm with the interior. (B) SEM micrograph taken with a JEOL JSM7600F of the 3D scaffold surface, showing mesoporous channels typical of a 2D hexagonal structure. (C) TEM image showing a mesoporous arrangement, where the darker zones homogeneously distributed into the mesoporous channels correspond to the apatite nanocrystalline phase. | ||
Moreover, these porosity properties can lead to mineralization, vascularisation and nutrient diffusion, which are critical for osteogenesis.27–29 Furthermore, the mesoporous arrangement could result in an added value as controlled delivery systems of biologically active molecules.30,31
With the aim of evaluating the in vitro degradation of MGHA 3D scaffolds, the surface changes and the evolution of their macroporous structure after different incubation times in the culture medium have been studied by SEM (Fig. 2). In the first 24 h, the SEM micrographs (Fig. 2D–F) show an early gradual degradation of the surface with small deposits of particles.
 | ||
| Fig. 2 In vitro degradation test. SEM micrographs at different magnifications and EDS analyses (inset) before (A–C) and after 1 (D–F) and 7 day (G–I) soaking in DMEM. | ||
Fig. 3 shows in detail this gradual deposition which is formed by needle-like particles typical of apatite formation on bioactive surfaces.29 On the other hand, after 7 days, there are substantial changes in the surface and the macroporous architecture of the scaffold (Fig. 2G and H), showing a notable degradation from a smooth surface (in situ) to a highly degraded surface which exhibits macropores in the range of 100–25 μm. Moreover, the higher magnification SEM image (Fig. 2I) shows the existence in this surface of acicular particles with a majority of Ca and P (Ca/P = 1.67), corresponding to the apatite phase, in agreement with other studies.32 EDS analyses (insets in Fig. 2C, F and I) confirm both the notable increase of the amount of Ca and P together with a decrease of Si amount in the scaffold surface after 1 and 7 days of incubation in DMEM, respectively (Fig. 4). These results evidence the excellent bioactive behaviour, according to previous results in simulated body fluid.9
 | ||
| Fig. 3 SEM study of MGHA scaffolds before and after 24 hours of incubation in DMEM. The micrographs show clearly the formation of needle-like aggregates of apatite phase on the scaffold surface after 24 h. | ||
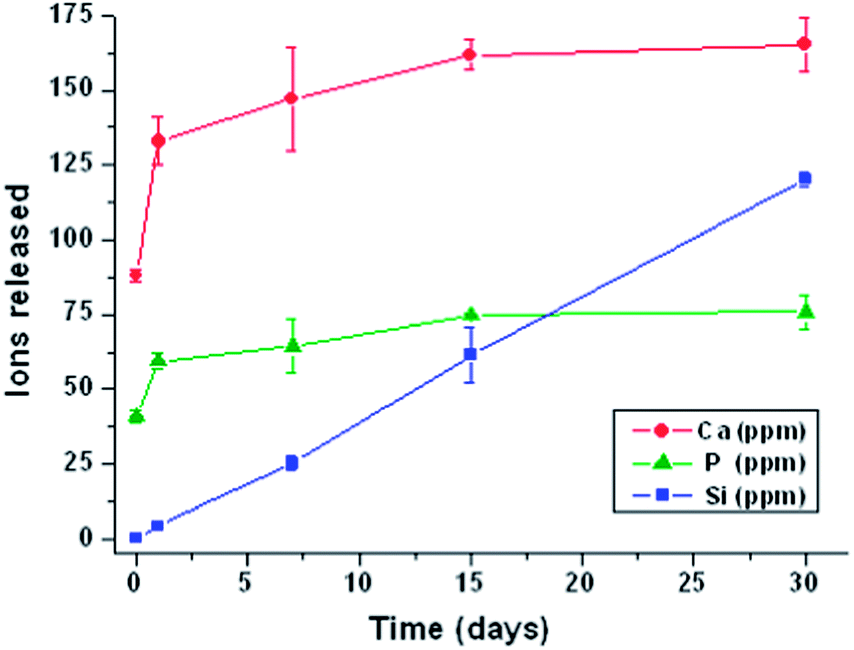 | ||
| Fig. 4 Variation of the calcium, phosphorous and silicon concentrations in DMEM over time. | ||
Fig. 4 shows a gradual degradation of 3D scaffolds, exhibiting an initial rapid increase of calcium and phosphorus concentrations in the soaking medium in the first 24 h of incubation. Then, the ionic concentrations slightly increase until 7 days and remain without variation until 30 days. Concerning the silicon variation, the plot shows constant gradual silicon leaching during all time periods. Thus, the obtained results reveal a gradual degradation of these 3D MGHA scaffolds over time.
FTIR spectroscopy was carried out to evaluate the changes in the MGHA nanocomposite scaffold surface as a function of soaking time in DMEM (Fig. 5). Before soaking, the spectrum shows characteristic absorption bands of Si–O bonds at 1040, 800, and 470 cm−1. After 24 hours of soaking in DMEM, an increase of the doublet at 560 and 600 cm−1, corresponding to a crystalline phosphate, is observed, confirming the formation of a new crystalline phosphate onto the scaffold surface which is in agreement with SEM-EDS results. Moreover, after 7 days, the spectrum analysis shows clearly the protein adsorption derived of complete DMEM onto the MGHA surface formed onto the new apatite layer.
 | ||
| Fig. 5 FTIR spectra corresponding to MGHA scaffolds before and after 24 h and 7 days in DMEM. | ||
To evaluate in vitro the response of immune cells to MGHA scaffolds, RAW-264.7 macrophage-like cells and murine spleen cells have been cultured in contact with these samples or their extracts (obtained after 24 h and 7 days as described above in Section 2.3). These incubation times have been chosen taking into account the higher reactivity of the MGHA material at 24 h and the scaffold degradation/bioactivity observed after 7 days.
3.2. Effects of the MGHA biomaterial on RAW-264.7 macrophage-like cells
RAW-264.7 macrophage-like cells are functional macrophages with the ability of autophagy and phagocytosis as immune cells and retain many of the characteristics of macrophages in vivo; for these reasons this cell line is usually used for in vitro biocompatibility studies.33Table 1 shows that no significant changes of cell viability (up 75%) were detected by the propidium iodide exclusion test after 24 h culture of RAW-264.7 macrophage-like cells with MGHA extracts (obtained at 24 h and 7 days; E 24 h and E 7 d, respectively) or MGHA scaffolds (SC).| Sample | Viability (%) |
|---|---|
| Control | 79.3 ± 2.2 |
| E 24 h | 78.8 ± 2.8 |
| E 7 d | 82.1 ± 3.4 |
| SC | 75.8 ± 2.1 |
When proliferation of RAW-264.7 macrophage-like cells on MGHA scaffolds was evaluated, no alterations were detected in comparison with control samples (Table 2). The cell cycle was studied by flow cytometry after culture of these cells on MGHA scaffolds and the SubG1 fraction, attributed to cells with fragmented DNA, was used as an indicator of apoptosis.34 A viable cell number count was used to assess proliferation. As it can be observed in Table 2, MGHA samples did not produce changes in the SubG1 phase of RAW-264.7 macrophage-like cells, thus indicating that MGHA scaffolds did not induce apoptosis in this cell type which presented apoptosis levels lower than 3%.
| Sample | Proliferation (cell number × 10−3) | Apoptosis (%) |
|---|---|---|
| Control | 32.2 ± 6.2 | 2.81 ± 0.14 |
| SC | 30.5 ± 4.4 | 2.57 ± 0.13 |
When confocal microscopy techniques were carried out to evaluate the RAW-264.7 morphology in contact with MGHA disks, the typical characteristics of these macrophages were observed. Fig. 6 shows that no morphological alterations induced by the MGHA material were detected in RAW-264.7 macrophage-like cells which showed intact cytoplasmic filaments of F-actin (Fig. 6, FITC phalloidin is labeled in green). DAPI is a DNA dye widely used to visualize the changes of the chromatin in apoptotic cells. This probe stains the cell nuclei and allows detection of the presence of apoptotic bodies produced by chromatin condensation/fragmentation as a late marker of apoptosis. No apoptotic nuclei were detected in RAW-264.7 macrophage-like cells cultured on MGHA disks in agreement with the low levels of apoptosis detected by flow cytometry (Table 2).
 | ||
| Fig. 6 Morphological evaluation of RAW-264.7 macrophage-like cells on MGHA disks. Cells were stained with DAPI for the visualization of the cell nuclei in blue and with FITC phalloidin for the visualization of cytoplasmic F-actin filaments in green. | ||
The effects of 24 h and 7 day MGHA extracts and scaffolds on the cell size and complexity of RAW-264.7 macrophage-like cells were also evaluated by flow cytometry through FSC and 90° SSC light scatter as indicators of cell size and complexity, respectively. These properties are determined in part by cell size, plasma membrane, cytoplasm, mitochondria, pinocytic vesicles, and lysosomes.35 As it can be observed in Fig. 7, the contact of RAW-264.7 macrophage-like cells with MGHA scaffolds or their extracts produces a decrease of cell size (Fig. 7A) with a slight increase of macrophage complexity (Fig. 7B).
 | ||
| Fig. 7 Effect of MGHA extracts and MGHA scaffolds on light scattering properties of RAW-264.7 macrophage-like cells. (A) Forward angle scatter (FSC, cell size) versus number of cells after treatment with MGHA scaffolds (green), 24 h extract (pink), and 7 day extract (blue). (B) 90 side angle light scatter (SSC, internal complexity) versus number of cells after treatment with MGHA scaffolds (green), 24 h extract (pink), and 7 day extract (blue). Control cells without treatment (black). | ||
Macrophages are one of the most versatile types of immune cells, carrying out a large number of important functions, essential to both innate and acquired immune responses. These functions include: phagocytosis of apoptotic cells, bacteria, and viruses; production of reactive nitrogen and oxygen species; antigen processing and presentation; and cytokine and chemokine production.15 Since reactive oxygen species (ROS) and Ca2+ constitute the most important intracellular signaling molecules for the regulation and integration of diverse cellular functions, the effects of MGHA extracts (obtained at 24 h and 7 days) and MGHA scaffolds (SC) on the intracellular ROS content and cytosolic calcium of RAW-264.7 macrophage-like cells after 24 h treatment were evaluated by flow cytometry with the probes DCFH and Indo-1 respectively. Significant ROS increases (p < 0.005) were produced by 7 day MGHA extracts and scaffolds (Table 3, a.u. = arbitrary units of fluorescence). Concerning intracellular calcium measurements, the probe Indo-1 shifts their spectral characteristics upon calcium binding, from FL1 to FL2;36 the ratio FL1/FL2 can be used as an indicator of the intracellular Ca2+ content. As it can be observed in Table 3, the fluorescence ratio of RAW-264.7 macrophage-like cells significantly increased (p < 0.001) after culture with 24 h and 7 day MGHA extracts and scaffolds, thus indicating that the MGHA extracts and scaffolds induced a significant cytosolic calcium increase in these cells. Several studies have demonstrated that some materials increase [Ca2+]i possibly through the entry of extracellular Ca2+via Ca2+ channels in the plasma membrane, and activate the generation of ROS.37
| Sample | ROS (a.u.) | Cai (FL1/FL2) |
|---|---|---|
| Control | 476 ± 23.8 | 1.70 ± 0.09 |
| E 24 h | 471 ± 23.6 | 2.47 ± 0.12 |
| E 7 d | 606 ± 30.3 | 2.36 ± 0.12 |
| SC | 611 ± 30.6 | 2.45 ± 0.12 |
Macrophages play a central role in directing the host response to implanted biomaterials, including the inflammatory response and the reparative response with M1 and M2 phenotypic spectra.17–22 Since several studies have demonstrated that M1 macrophages produce and secrete higher ROS levels than M2 cells,17,38 the effects of MGHA extracts or MGHA scaffolds on populations of RAW-264.7 macrophage-like cells with low and high intracellular ROS contents have been evaluated by flow cytometry in the present study. Fig. 8 shows the profiles of cell number (counts) versus DCF fluorescence (FL1) used as an indicator of intracellular ROS. Macrophages in contact with MGHA scaffolds (Fig. 8B) and control macrophages (Fig. 8A) showed different intracellular ROS profiles. Fig. 9 shows that in the presence of 24 h extracts, the population with a low ROS content is significantly higher than the population with a high ROS content, in agreement with the results presented in Table 3 which indicates no high intracellular ROS levels when RAW macrophage-like cells are cultured with these extracts. This effect can be related to the rapid release of ions (calcium, phosphorous and silicon) to the medium in the early steps of the reactivity/bioactivity process (Fig. 4). On the other hand, the 7 day MGHA extract does not induce changes in the percentages of low and high ROS populations probably due to the stabilization of this release which is slower at this time (Fig. 4). The direct contact of RAW-264.7 macrophage-like cells with the MGHA scaffolds for 24 h increases both the intracellular ROS content (Table 3) and, subsequently, the high ROS content population in comparison with control, thus indicating the high reactivity/bioactivity of the surface-level of MGHA scaffolds previously observed in Fig. 3 and 4.
 | ||
| Fig. 8 Effect of MGHA scaffolds on populations of RAW-264.7 macrophage-like cells evaluated through intracellular ROS (flow cytometry profiles). Intracellular ROS (FL1, a.u.) versus number of cells after treatment with MGHA scaffolds (B). Control untreated cells (A). | ||
 | ||
| Fig. 9 Effect of MGHA extracts and MGHA scaffolds on the percentage of populations of RAW-264.7 macrophage-like cells with low and high intracellular ROS contents. CTL = control untreated cells. Columns with different letters are statistically different. Statistical significance **ρ < 0.01; ***ρ < 0.005. | ||
3.3. Effects of MGHA biomaterial on murine spleen cell suspensions
Immune response plays a determinant role in the tolerance to and functioning of biomaterials. The immune system is composed of many different subpopulations which mutually interact and cross-talk determining the final outcome of the immune response. To assess the potential effect of MGHA on immune cells, we chose to analyze its effect on murine spleen cell suspensions, an in vitro system in which different untransformed normal cell subpopulations interact among them, better resembling the in vivo ambient environment. The spleen is a lymphoid organ which contains different types of immune cells, mainly including T- and B-lymphocytes (about 80% in the mouse spleen), macrophages and other antigen-presenting (APC) (about 10%) and natural killer (NK) cells. These cells circulate through the body monitoring for signals which trigger an innate response or detecting non-self antigens, thus becoming activated, differentiated and expanded, mounting an acquired immune response. The contact of body fluids and blood cells with biomaterials may trigger the activation of granulocytes, monocytes and macrophages, driving to an acute and late chronic inflammatory response. Although the biomaterial composition does not allow T cell activation via antigen presentation, it has been suggested that synthetic biomaterials may present functional groups on their surfaces acting as mitogens which can trigger lymphocytes by cross-linking glycoproteins on the cell surface.15 We assessed this possibility by studying the effect of MGHA extracts and scaffolds on murine normal and activated immune cells.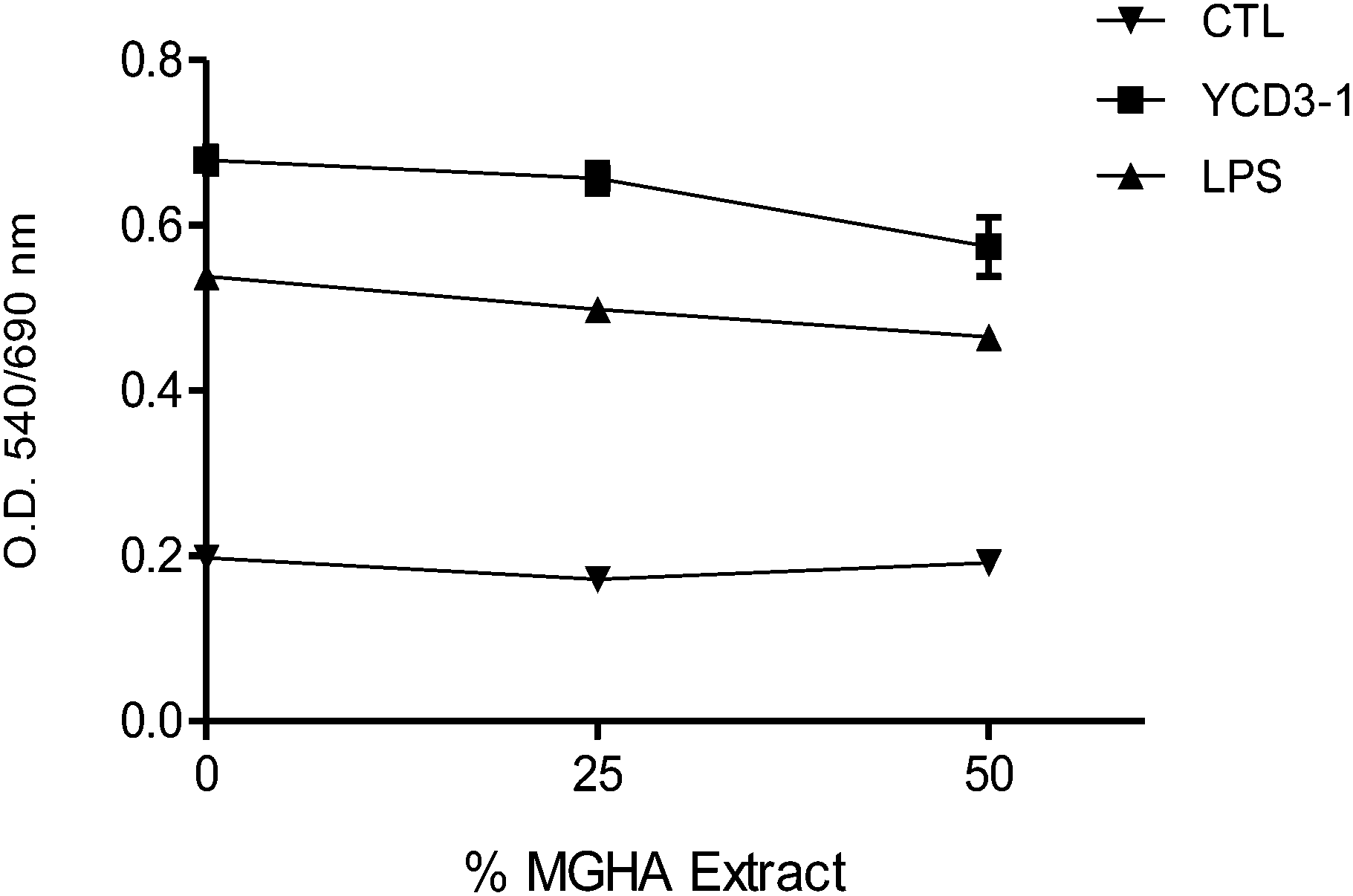 | ||
| Fig. 10 Effect of MGHA extracts on murine spleen cell activation. The effect of two different concentrations of the MGHA extract was evaluated in an activation assay in the presence of anti-CD3 antibody (YCD3-1, T-cell mitogen, squares), LPS (B-cell mitogen, triangles) or medium (control, CTL, inverted triangle). Mitochondrial activity was measured after 72 hour culture and represented as O.D. | ||
Fig. 11 shows that no significant modification of spontaneous apoptosis (p > 0.05 in all cases) was induced by MGHA scaffolds in T (CD3+, CD4+, and CD8+) or B (CD19+) lymphocytes, natural killer (NK1.1+, GR1+, and CD11b+) or monocytes–macrophages (CD11b+) subpopulations in the spleen cell cultures. GR1 is also a marker for the low proportion of peripheral granulocytes found in the spleen which was not affected by MGHA in this assay.
 | ||
| Fig. 11 MGHA scaffolds do not affect spontaneous apoptosis of different spleen cell subpopulations. Apoptosis was measured as a percent of Annexin V+propidium iodide− cells by FACS in murine spleen cells which were seeded onto MGHA scaffolds (filled bars) or directly in the culture wells (CTL, empty bars) and cultured overnight. | ||
 | ||
| Fig. 12 MGHA scaffolds do not affect the subpopulations in a murine spleen cell activation assay. The percentage of different spleen cell subpopulations (indicated in the X axis) was evaluated by FACS analysis after 72 hour culture under T- (anti-CD3 antibody) (A) or B-cell stimulus (LPS) (B) in the presence (filled bars) or absence (CTL, empty bars) of MGHA scaffolds. | ||
Cytokine expression and secretion constitute an important effector mechanism of immune cells, determining subsequent activation or suppression of other subpopulations. Among them, we chose to analyse IL-2, IL-10, IFNγ and TNFα because of their important implication in the immune response and inflammation. IL-2 is a Th1 cytokine that plays a central role in the activation, proliferation and differentiation of Ag-primed T lymphocytes.40,41 It also modulates the expression of IFNγ and MHC antigens, stimulates proliferation and differentiation of activated B cells and augments NK cell activity. IL-10 is an anti-inflammatory Th2 cytokine that has a critical role in limiting the immune response to prevent host damage.42 It is expressed by Th2, Th17, T regulatory and B cells, as well as innate immune cells as dendritic cells, macrophages, mast cells, NK, eosinophils and neutrophils. As IL-10 is produced in several T helper populations, it is proposed that it provides a feedback loop to limit the effector function of macrophages and dendritic cells. IFN-γ (Type-II Interferon) is produced by activated T, B and NK cells. During infection it is produced by cytotoxic CD8+ T cells and Th1, inhibiting the proliferation of Th2 phenotype subpopulation. IFNγ not only acts in synergy with other cytokines including TNFα to inhibit the proliferation of normal or transformed cells but also activates monocytes, macrophages, NK and neutrophils. IFNγ is a marker for a number of different pathological situations including infections, autoimmune diseases, transplant rejection and diabetes.43,44 TNF-α (Tumor Necrosis Factor alpha) is a pleiotropic cytokine that plays a role in innate and adaptive immunity.45 Though it is widely implicated in numerous immune responses and regulations, it is most often associated with regulation of cell survival and pro-inflammatory properties which lead to the recruitment and activation of inflammatory cells to the site of injury where it is known to induce various cytokines, including IL-1, IL-6, IL-8, MCP-1 and RANTES. TNF-α is primarily expressed by macrophages, but also by activated lymphocytes, NK cells and neutrophils, among others. It is associated with numerous inflammatory illnesses and autoimmunity. To analyze the potential secondary effect of MGHA on immune cells we studied the intracellular expression of IL-2, IL-10, IFN-γ or TNF-α in spleen cell cultures which were activated by mitogens for 72 h. Cytokine expression was analysed in total spleen cells or CD3+ or CD19+ subpopulations. No significant effect (p > 0.05) of MGHA scaffolds on the intracellular cytokine expression was detected in the analysed cell populations under T- (Fig. 13A) or APC-activating conditions (Fig. 13B). The absence of significant changes in IL-2 and IFNγ expression in the presence of the scaffolds indicates the lack of MGHA effects on T cells (Fig. 13). In parallel, the absence of significant changes in TNF-α or IL-10 expression indicates that pro- or anti-inflammatory loops are not significantly modified by MGHA scaffolds under T- or APC-stimuli conditions (Fig. 13A and B).
 | ||
| Fig. 13 MGHA biomaterial does not affect the cytokine expression in activated murine immune cells. The percent of cytokine positive spleen cells (indicated in the X axis) was evaluated by FACS analysis after 72 hour culture under T- (anti-CD3 antibody) (A) or B-cell stimulus (LPS) (B) in the presence (filled bars) or absence (CTL, empty bars) of MGHA scaffolds. | ||
Adjuvant properties of mesoporous silica nanoparticles have been recently shown in several experimental models, producing different effects which depend on the size and functionalization of the particles.46–48 In this sense, amino-functionalized MCM-41 (AM-41) has been used as a vaccine delivery vehicle and an adjuvant in mice using ovalbumin as a model protein antigen.48 AM-41 nanoparticles showed adjuvant behaviour and elicited immune responses at reduced antigen doses in vivo. However, there were no local or systemic negative effects on animals injected with AM-41.48 In the present study, MGHA scaffolds do not stimulate a significant inflammatory response in vitro. The absence of MGHA immunogenic activity might be explained considering the morphology of the samples (which are scaffolds and not nanoparticles) and the different evaluation models.
4. Conclusions
Some modifications are induced by the MGHA nanocomposite on RAW macrophage-like cells after contact with the scaffold surface or with 24 h extracts, which can be related to the high reactivity/bioactivity of this nanocomposite. However, the MGHA material does not produce significant in vitro alterations in the activation of macrophages, B- or T-lymphocytes and natural killer cells when an immune response stimulation model was carried out using murine primary immune cell subpopulations. Since the MGHA scaffolds are good supports for adhesion, proliferation and differentiation of preosteoblasts, and they would not stimulate a significant inflammatory response, the present study supports the biocompatibility of the MGHA material and suggests an adequate host tissue response to their scaffolds upon their implantation.Acknowledgements
This study was supported by research grants from Comunidad de Madrid (S2009/MAT-1472), and Ministerio de Economía y Competitividad (MINECO) (research grants MAT2012-35556, CS2010-11384-E and FIS PI10/0648). Mónica Cicuéndez is grateful to MICINN for the financial support through FPI fellowship. The authors wish to thank also the staff of the ICTS Centro Nacional de Microscopia Electrónica (Spain) and Centro de Citometría y Microscopia de Fluorescencia. The authors thank Dr Shunsuke Asahina (SMBU, JEOL, Akishima, Tokyo 196-8558, Japan) and Prof. Osamu Terasaki (Graduate School of EEWS, WCU, KAIST, Daejeon, Republic of Korea) for their help in tuning the JEOL JSM7600F microscope in the ICTS (http://www.cnme.es) and capturing the micrographs.Notes and references
- R. Langer and J. P. Vacanti, Science, 1993, 260, 920–926 CAS.
- T. G. Kim, H. Shin and D. W. Lim, Adv. Funct. Mater., 2012, 22, 2446–2468 CrossRef CAS.
- F. J. O'Brien, Mater. Today, 2011, 14, 88–95 CrossRef CAS.
- S. K. Misra, T. I. Ansari, S. P. Valappil, D. Mohn, S. E. Philip, W. J. Stark, I. Roy, J. C. Knowles, V. Salih and A. R. Boccaccini, Biomaterials, 2010, 31, 2806–2815 CrossRef CAS PubMed.
- X. Wang, G. Song and T. Lou, Med. Eng. Phys., 2010, 32, 391–397 CrossRef PubMed.
- M. T. Arafat, C. X. F. Lam, A. K. Ekaputra, S. Y. Wong, X. Li and I. Gibson, Acta Biomater., 2011, 7, 809–820 CrossRef CAS PubMed.
- M. Ebrahimian-Hosseinabadi, F. Ashrafizadeh, M. Etemadifar and S. S. Venkatraman, Polym. Degrad. Stab., 2011, 96, 1940–1946 CrossRef CAS PubMed.
- X. Y. Lu, T. Qiu, X. F. Wang, M. Zhang, X. L. Gao, R. X. Li, X. Lu and J. Weng, Appl. Surf. Sci., 2012, 262, 227–230 CrossRef CAS PubMed.
- M. Cicuéndez, M. T. Portolés, I. Izquierdo-Barba and M. Vallet-Regí, Chem. Mater., 2012, 24, 1100–1106 CrossRef.
- M. Alcaide, M. T. Portolés, A. López-Noriega, D. Arcos and M. Vallet-Regí, Acta Biomater., 2010, 6, 892–899 CrossRef CAS PubMed.
- M. Cicuéndez, M. Malmsten, J. C. Doadrio, M. T. Portolés, I. Izquierdo-Barba and M. Vallet-Regí, J. Mater. Chem. B, 2014, 2, 49–58 RSC.
- S. Hollister, Adv. Mater., 2009, 21, 3330–3342 CrossRef CAS PubMed.
- D. W. Hutmacher, Biomaterials, 2000, 21, 2529–2543 CrossRef CAS.
- A. Remes and D. F. Williams, Biomaterials, 1992, 13, 731–743 CrossRef CAS.
- S. Franz, S. Rammelt, D. Scharnweber and J. C. Simon, Biomaterials, 2011, 32, 6692–6709 CrossRef CAS PubMed.
- M. J. Smith, K. L. White, D. C. Smith and G. L. Bowlin, Biomaterials, 2009, 30, 149–159 CrossRef CAS PubMed.
- B. N. Brown, B. D. Ratner, S. D. Goodman, S. Amar and S. F. Badylak, Biomaterials, 2012, 33, 3792–3802 CrossRef CAS PubMed.
- D. M. Mosser and J. P. Edwards, Nat. Rev. Immunol., 2008, 8, 958–969 CrossRef CAS PubMed.
- B. N. Brown, R. Londono, S. Tottey, L. Zhang, K. A. Kukla, M. T. Wolf, K. A. Daly, J. E. Reing and S. F. Badylak, Acta Biomater., 2012, 8, 978–987 CrossRef CAS PubMed.
- P. M. Kou and J. E. Babensee, J. Biomed. Mater. Res., Part A, 2010, 96A, 239–260 CrossRef PubMed.
- K. Garg, N. A. Pullen, C. A. Oskeritzian, J. J. Ryan and G. L. Bowlin, Biomaterials, 2013, 34, 4439–4451 CrossRef CAS PubMed.
- R. M. Boehler, J. G. Graham and L. D. Shea, Biotechnology, 2011, 51, 239–254 CAS.
- P. Portolés, J. M. Rojo, A. Golby, M. Bonneville, S. Gromkowski, L. Greenbaum, C. A. Janeway and D. B. Murphy, J. Immunol., 1989, 142, 4169–4175 Search PubMed.
- T. Mosmann, J. Immunol. Methods, 1983, 65, 55–63 CrossRef CAS.
- J. M. Rojo, E. Pini, G. Ojeda, R. Bello, C. Dong, R. A. Flavell, U. Dianzani and P. Portolés, Int. Immunol., 2008, 20, 577–589 CrossRef CAS PubMed.
- M. Cicuéndez, I. Izquierdo-Barba, M. T. Portolés and M. Vallet-Regí, Eur. J. Pharm. Biopharm., 2013, 84, 115–124 CrossRef PubMed.
- M. Vallet-Regí and J. M. González-Calbet, Prog. Solid State Chem., 2004, 3, 1–31 CrossRef PubMed.
- J. R. Jones, Acta Biomater., 2013, 9, 4457–4486 CrossRef CAS PubMed.
- M. Vallet-Regí, M. Colilla and I. Izquierdo-Barba, J. Biomed. Nanotechnol., 2008, 4, 1–15 CrossRef PubMed.
- M. Vallet-Regí, ISRN Mater. Sci., 2012, 608548 Search PubMed , 20 pages.
- M. Vallet-Regí, J. Intern. Med., 2010, 267, 22–43 CrossRef PubMed.
- M. Vallet-Regí, C. V. Ragel and A. J. Salinas, Eur. J. Inorg. Chem., 2003, 6, 1029–1042 CrossRef.
- H. Kim, B. Cheon, Y. H. Kim, S. Y. Kim and H. P. Kim, Biochem. Pharmacol., 1999, 58, 759–765 CrossRef CAS.
- M. Alcaide, M. C. Serrano, R. Pagani, S. Sánchez-Salcedo, M. Vallet-Regí and M. T. Portolés, Biomaterials, 2009, 30, 45–51 CrossRef CAS PubMed.
- J. M. Udall, R. A. Moscicki, F. I. Preffer, P. D. Ariniello, E. A. Carter, A. K. Bhan and K. J. Bloch, Adv. Exp. Med. Biol., 1987, 216, 821–827 CrossRef.
- G. Grynkiewicz, M. Poenie and R. Y. Tsien, J. Biol. Chem., 1985, 260, 3440–3450 CAS.
- F. Tian, T. Zhu and Y. Shang, Toxicol. Appl. Pharmacol., 2010, 242, 173–181 CrossRef CAS PubMed.
- S. Franz, F. Allenstein, J. Kajahn, I. Forstreuter, V. Hintze and S. Möller, Acta Biomater., 2013, 9, 5621–5629 CrossRef CAS PubMed.
- W. G. Brodbeck, M. Macewan, E. Colton, H. Meyerson and J. M. Anderson, J. Biomed. Mater. Res., Part A, 2005, 74A, 222–229 CrossRef CAS PubMed.
- S. L. Gaffen and K. D. Liu, Cytokine, 2004, 28, 109–123 CrossRef CAS PubMed.
- J. Banchereau, V. Pascual and A. O'Garra, Nat. Immunol., 2012, 13, 925–931 CrossRef CAS PubMed.
- M. Saraiva and A. O'Garra, Nat. Rev. Immunol., 2010, 10, 170–181 CrossRef CAS PubMed.
- K. Schroder, P. J. Hertzog, T. Ravasi and D. A. Hume, J. Leukocyte Biol., 2004, 75, 163–189 CrossRef CAS PubMed.
- L. C. Platanias, Nat. Rev. Immunol., 2005, 5, 375–386 CrossRef CAS PubMed.
- M. Croft, Nat. Rev. Immunol., 2009, 9, 271–285 CrossRef CAS PubMed.
- H. Vallhov, N. Kupferschmidt, S. Gabrielsson, S. Paulie, M. Strømme, A. E. Garcia-Bennett and A. Scheynius, Small, 2012, 8, 2116–2124 CrossRef CAS PubMed.
- X. Wang, X. Li, A. Ito, Y. Sogo and T. Ohno, Acta Biomater., 2013, 9, 7480–7489 CrossRef CAS PubMed.
- D. Mahony, A. S. Cavallaro, F. Stahr, T. J. Mahony, S. Z. Qiao and N. Mitter, Small, 2013, 9, 3138–3146 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2014 |