
Crystal structure and substrate-binding mode of the mycoestrogen-detoxifying lactonase ZHD from Clonostachys rosea†
Abstract
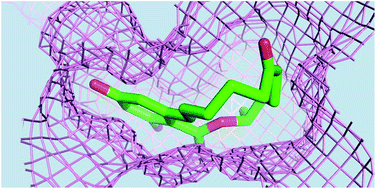
The mycotoxin zearalenone has been contaminating maize and other grains. It can be hydrolyzed and inactivated by the lactonase ZHD, which belongs to the α/β-hydrolase family. Besides the catalytic core domain, the enzyme comprises an α-helical cap domain. Zearalenone differs from other quorum-sensing lactones in its chemical structure. As revealed by the complex structure, the substrate binds into a deep pocket between the core and cap domains, adjacent to the catalytic triad Ser102–His242–Glu126. The enzyme–substrate interactions include three direct hydrogen bonds and several nonpolar contacts. In particular, the Trp183 side chain is engaged in both hydrogen bonding and T-stacking interactions with the benzoate ring. The central role of Trp183 in substrate binding was verified by the mutants W183A, W183H and W183F. Several mutants were also produced to investigate the roles of nearby amino-acid residues. Interestingly, mutants that destabilize the dimer had adverse functional effects on ZHD.
 Please wait while we load your content...
Please wait while we load your content...

