
Filipins: the first antifungal “weed killers” identified from bacteria isolated from the trap-ant†
Abstract
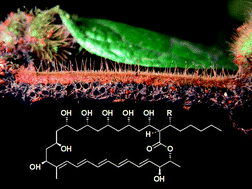
Allomerus ants ensure that they have sufficient nitrogen in their diet by trapping and consuming other insects. In order to construct their traps, like the more extensively studied leaf cutter ants, they employ fungal farming. Pest management within these fungal cultures has been speculated to be due to the ants' usage of actinomycetes capable of producing antifungal compounds, analogous to the leafcutter ant mutualism. Here we report the first identification of a series of antifungal compounds, the filipins, and their associated biosynthetic genes isolated from a bacterium associated with this system.
 Please wait while we load your content...
Please wait while we load your content...

