
Antioxidant and hepatoprotective effects of an organic grapevine leaf (Vitis labrusca L.) extract in diabetic rats
Abstract
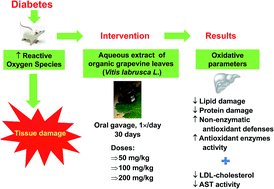
Our objective was to investigate the antioxidant effect of an aqueous extract of organic grapevine leaves (Vitis labrusca L.) on the livers of diabetic rats and to evaluate the resulting changes in metabolic and biochemical parameters. Diabetic rats received daily intragastric doses of 50, 100 or 200 mg kg−1 of the grapevine extract for 30 days. Grapevine leaf extract showed a dose-dependent antioxidant effect on the livers of diabetic rats, evidenced by decreases in TBARS and in carbonyl levels and increases in sulfhydryl levels. Moreover, the extract (200 mg kg−1) prevented weight loss and reduced LDL cholesterol (50 mg kg−1), urea (50 mg kg−1), and AST (50 and 100 mg kg−1) levels in diabetic rats at the indicated doses. Thus, we suggest that chronic treatment with an extract of grapevine leaves may represent an adjuvant therapy for the treatment and/or prevention of diabetic complications because of its antioxidant, hepatoprotective and possible hypolipidemic effects shown here.
 Please wait while we load your content...
Please wait while we load your content...

