
Green remediation of textile dyes containing wastewater by Ipomoea hederifolia L.†
Niraj R. Ranea,
Vishal V. Chandanshiveb,
Rahul V. Khandarec,
Avinash R. Gholaved,
Shrirang R. Yadavd and
Sanjay P. Govindwar*b
aDepartment of Biotechnology, Shivaji University, Kolhapur, India
bDepartment of Biochemistry, Shivaji University, Kolhapur, India. E-mail: spg_biochem@unishivaji.ac.in; Fax: +91-231-2691533; Tel: +91-231-2609152
cSchool of Nanoscience and Technology, Shivaji University, Kolhapur, India
dDepartment of Botany, Shivaji University, Kolhapur, India
First published on 24th July 2014
Abstract
Wild plant and tissue cultures of Ipomoea hederifolia decolorize Scarlet RR (SRR) dye at a concentration of 50 mg L−1 up to 96% and 90% within 60 and 96 h, respectively. Significant induction in the enzyme activities of Lignin peroxidase, laccase, 2,6-dichlorophenol indophenol reductase, superoxide dismutase, catalase and tyrosinase was found in the plant roots and shoots during decolorization. I. hederifolia was also able to treat a dye mixture and a real textile effluent efficiently with a reduction in the American Dye Manufacturers Institute (ADMI) value (color removal) up to 85% and 88%, BOD up to 65% and 63% and COD up to 62% and 68%, respectively. Detailed anatomical studies of the stem and root cells of I. hederifolia during uptake and degradation were carried out, showing a stepwise and mechanistic degradation of the model dye SRR. Products formed after dye degradation were analyzed by UV-Vis spectroscopy, FTIR, HPLC and HPTLC, which confirmed the phytotransformation of SRR, dye mixture and textile effluent. A possible pathway for the phytotransformation of SRR was proposed based on GC-MS analysis, which confirmed the formation of different metabolites with lower molecular weights. The phytotoxicity study revealed the non-toxic nature of the formed products.
1. Introduction
Industrial developments are always associated with a cost in terms of pollution of air, water bodies and soil by a number of toxic compounds. The fast growing textile industry is one of the sources to release toxic chemical compounds in the form of dyes. Note that approximately 10–15% of the synthetic dyes having carcinogenic and mutagenic effects are released during the dying process, ultimately threatening all life forms.1 The valuable water bodies needed for irrigation, drinking and other household and industrial purposes are being polluted because of the textile dyes released through effluent. Inclusion of textile dyes in water reservoirs alters the vital parameters of water bodies by influencing the levels of chemical oxygen demand (COD), biological oxygen demand (BOD), total organic carbon (TOC), total dissolved solids (TDS), total suspended solids (TSS), pH and color.2 Therefore, addressing the issue of these environmental contaminants has become very important and needs attention from environmentalists.3 This, at the end, results in the development of odour and deterioration of overall water quality in the reservoir. The ability of textile dyes to remain recalcitrant to degradation in water and soil can be ascribed to their complex chemical structure. The persistence of toxic compounds in aquatic and soil environments may lead to accumulation in phytoplankton, fish and vegetables.4 In developing countries, the textile dyes are not only part of textile industry effluent but also add to the municipal sewages.3A number of physicochemical methods, such as adsorption, coagulation, flocculation, filtration, photodegradation and chemical oxidation, are available for dealing with the pollution created by textile dyes. These methods are associated with high cost, low efficiency, secondary pollution problems and the inability to treat a wide array of dyes having structural diversity.2 Although these methods are employed to remove the colour, their capacity to reduce the toxicity is still a matter of major concern. These facts certainly demand the development of an efficient, cost effective and eco-friendly technology for the removal of these dyes from environment. The use of microorganisms for purifying dye contaminated water has extensively been studied and demonstrated but is still associated with some limitations as far as in situ administration of pollutants is concerned.5
The use of plants to decolorize and detoxify the textile dyes in the laboratory, as well as at the actual site of contamination, appears as an attractive option.6 Phytoremediation appears to be an ideal potential approach for the treatment of pollutants; it is more admired because of its solar-driven nature, cost effectiveness, aesthetic advantages and long-standing applicability as it can be directly employed at the polluted sites compared to other expensive methods of treatment.7 Some wild plants like Phragmites australis, Blumea malcolmii, Typhonium flagelliforme, Rheum rabarbarum (rhubarb) and R. hydrolapatum have been proposed for textile dye removal.6,8–11 Recently, common garden plants such as Aster amellus, Glandularia pulchella, Portulaca grandiflora, Petunia grandiflora, Zinnia angustifolia and Tagetes patula have been shown to have great potential for textile dye degradation.12–18 Though the use of nursery-grown garden plants appears attractive, their use for the clean-up of actually contaminated sites has yet to be demonstrated.
This work deals with the decolorization and degradation of a commonly used disperse textile dye Scarlet RR (SRR), a dye mixture and a real textile effluent by Ipomea hederifolia Linn. This plant was procured from the actual site of dye disposal, Ichalkaranji, India. Therefore, it was thought to have the ability to grow in high dye concentrations; thus, it was better adapted to these dye stress conditions. The plant also possesses characteristics like a dense root system, the ability to grow in marshy places and a non-edible nature. Moreover, the seeds of I. hederifolia can germinate in the soil contaminated with dyes, which makes it a favourable candidate plant for phytoremediation of textile dyes. This work also explores the removal and degradation pattern by the plant at an anatomical level and reveals the histological dye degradation process by I. hederifolia.
2. Materials and methods
2.1. Chemicals
2,2′-Azino-bis(3-ethylbenzothiazoline)-6-sulphonic acid (ABTS) and riboflavin were available from Sigma Aldrich (St Louis, MO, USA). Nicotinamide adenine dinucleotide (di-sodium salt), n-propanol, 2,6-dichlorophenol indophenol (DCIP), veratryl alcohol and catechol were purchased from Sisco Research Laboratories, Mumbai, Maharashtra, India. Tartaric acid was available from BDH chemicals (Mumbai, Maharashtra, India). The MS Basal Medium was purchased from HiMedia Laboratories, Pvt. Ltd., India. The textile dyes SRR, Navy Blue HE2R, Green HE4B, Blue 2 RNL, Direct Red 5B, Reactive Red 2, Red HE3B, Remazol Red, Remazol Black B dyes and effluent were obtained from Mahesh, dye processors and Common Effluent Treatment Plant, Ichalkaranji, Maharashtra, India, respectively. All the chemicals used were of the highest purity available and an analytical grade. The seeds of Sorghum vulgare, Phaseolus mungo and Vigna radiata were purchased from a local grain market, and the seeds of I. hederifolia for tissue culture were purchased from Naamdhari Seeds Pvt. Ltd., India.2.2. Potential of I. hederifolia plants for dye decolorization
The plants of I. hederifolia were uprooted carefully and their roots were carefully washed with flowing water and exposed to 100 mL solutions of Navy Blue HE2R, Green HE4B, Blue 2 R, Direct Red 5B, Reactive Red 2, Red HE3B and SRR at a concentration of 50 mg L−1 independently. Based on the decolorization performance and longevity, I. hederifolia was selected for further studies.2.3. In vitro culture of I. hederifolia and its decolorization potential
The role of bacteria in the decolorization was doubted upon the first exposure to dye; therefore, in vitro grown cultures of I. hederifolia were obtained by a seed culture method. Seeds of I. hederifolia were collected from the plants found at the actual site of contamination, surface sterilized and cultured on MS basal medium as per a method previously described.5 The half-strength MS medium was discovered suitable for 90% of seed germination. A one week old seedling with profuse roots was used for phytoremediation studies. The tissue culture plants of I. hederifolia also showed degradation of SRR, mixture of textile dyes and real textile effluent taken for the study, which is explained below.2.4. Decolorization studies
Decolorization experiments were initially carried out with the wild plants in the dye SRR and the dye mixture containing Green HE4B, SRR, Remazol Black B, Remazol Red and Navy Blue HE2R. The structures and names of all the dyes used in this study are shown in the appendix. The experiments were carried out in 250 mL Erlenmeyer flasks containing 100 mL of 50 mg L−1 of dye solution in plain distilled water. As the bacterial contamination was doubted during decolorization, the experiments were independently performed with the tissue culturally grown plantlets of I. hederifolia exposing them to 15 mL of dye solution in test tubes. Absorbance of the dye solution was recorded at intervals of 6 h, each by removing 1 mL of solution. This solution was centrifuged at 4561g for 10 min to remove any solid matter if present and the absorbance of the clear solution was measured at a wavelength of 510 nm. The decolorization percentage was calculated as follows:
Abiotic controls contained the respective dye solutions, which were devoid of plants whereas plants in distilled water were kept as biotic controls.
2.5. Characterization of dye mixture and textile effluent
SRR, textile effluent and the mixture of dyes were characterized for American Dye Manufacturers Institute (ADMI) value,3 hardness, BOD, COD and TOC before and after dye degradation.192.6. Anatomical studies of stem and roots during dye degradation
Transverse sections of stem and root were taken and mounted in glycerin after overlaying with cover slip, and the results were micro-photographed with a Zeiss Axio Imager 2 Upright Trinocular Microscope with attached camera at 100× magnification.2.7. Photosynthetic pigments analysis
5 gm of leaves of both exposed and unexposed plants and a pinch of MgCO3 powder were crushed using a mortar and pestle, which contained 4 mL L−1 liquor ammonia. The extracts were centrifuged for 10 min at 2000g. Chlorophyll a, chlorophyll b and carotenoids were determined spectrophotometrically at 652, 645, and 663 nm.202.8. Preparation of cell free extract and enzyme assays
Roots and shoots of the plants of I. hederifolia were cut, weighed equally (2 g) were finely chopped and then suspended separately in 50 mM potassium phosphate buffer (pH 7.4). The chopped shoot tissue was then ground using a mortar and pestle, followed by homogenization in a glass homogenizer and then centrifugation at 8481g for 20 min. The cell free extract thus obtained was used as an enzyme source. The supernatant obtained after harvesting the plant roots and shoots was used as a source of extracellular enzymes after centrifugation. The cell-free solution obtained after harvesting cells was used as a source of extracellular enzymes.21Activities of the enzymes lignin peroxidase (LiP), veratryl alcohol oxidase, laccase, tyrosinase, DCIP reductase and azo reductase were determined spectrophotometrically at room temperature in the case of both the control and the test plants. LiP activity was determined by monitoring the formation of propanaldehyde at 300 nm in a reaction mixture of 2.5 mL containing 100 mM n-propanol, 250 mM tartaric acid, and 10 mM H2O2.22 Laccase activity was determined in a reaction mixture of 2 mL containing 10% ABTS in 0.1 M acetate buffer (pH 4.9), and the increase in optical density was measured at 420 nm.23 Tyrosinase activity was determined as described by an earlier report.24 NADH-DCIP reductase was measured in cell-free extract as reported earlier by Salokhe and Govindwar.25 Veratryl alcohol oxidase activity was determined by using veratryl alcohol as a substrate. The reaction mixture contained 1 mM veratryl alcohol, in 0.05 M citrate phosphate buffer (pH 3.0) and enzyme in a total volume of 2 mL, which was used for the determination of oxidase activity. Oxidation of the substrate at room temperature was monitored by observing an absorbance increase at 310 nm due to the formation of veratraldehyde. One unit of enzyme activity was measured as the amount of enzyme that releases 1 μmol product min−1.26 Antioxidant enzyme status was assessed by spectrophotometric assays. Antioxidant enzymes that were analyzed include catalase and superoxide dismutase.27
All enzyme assays were performed at 27 °C with reference blanks that contained all components except the enzyme. The protein contents of all the samples were determined using Lowry's method.28 All enzyme assays were run in triplicate, average rates were calculated and one unit of enzyme activity was defined as a change in absorbance unit min−1 mg of protein−1.
2.9. Analysis of the degradation products
Decolorization of all the dyes was monitored using UV-Vis spectroscopy analysis (Hitachi U-2800; Hitachi, Tokyo, Japan) using supernatants, whereas biotransformation was monitored using Gas Chromatography-Mass Spectrometry (GC-MS), High Performance Liquid Chromatography (HPLC), High Performance Thin Layer Chromatography (HPTLC) and Fourier Transform Infrared Spectroscopy (FTIR). For the extraction of products, plants were removed from the distilled water containing decolorized dye, which was then centrifuged to remove any solid matter (e.g., root hairs), if present. The decolorized and degraded solution was then extracted with equal volumes of ethyl acetate in a separating funnel. The organic extracts were combined and then evaporated in vacuum over anhydrous Na2SO4 and dried out. The solid residue obtained was dissolved in small volumes of HPLC grade methanol and these samples were used for analytical study.Identification of the metabolites produced was carried out using GC-MS. GC-MS analysis of the metabolites was carried out using a Shimadzu 2010 MS Engine equipped with an integrated gas chromatograph with an HP1 column (60 m long and 0.25 mm). Helium was used as the carrier gas at a flow rate of 1 mL min−1. The injector temperature was maintained by an oven at 80 °C for 2 min. The temperature was increased to 200 °C with a rate of 10 °C min−1 and then raised to 280 °C at a rate of 20 °C min−1. The compounds were identified on the basis of mass spectra and using the database of the National Institute of Structure and Technology (NIST) library. HPLC analysis was carried out (Waters model no. 2690; Waters Corp., Milford, MA) on a C18 column (symmetry, 4.6 mm × 250 mm) by using methanol with flow rate of 1 mL min−1 for 10 min and a UV detector at 254 nm. HPTLC analysis was carried out by using an HPTLC system (CAMAG, Switzerland). Samples of dye SRR, dye mixture and its biodegradation metabolites (dissolved in HPLC-grade methanol) were loaded on precoated HPTLC plates (Silica gel 60F 254, Merck, Germany), by using nitrogen as a spraying gas and a TLC sample loading instrument (CAMAG LINOMAT 5). The bands (12 mm) were applied at 10 mm from the lower edge of the plate with the first application position at 20 mm from the left edge of the plate and 5 mm apart from each other. The HPTLC plate was kept in a presaturated twin-trough chamber (10 × 20 cm) for the development of dyes and ethyl acetate extracted metabolites (products) with 10 mL of a standardized developing solvent system of toluene to methanol (8![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :2). After development, the plate was observed in a UV chamber (CAMAG) and scanned at 254 nm with a slit dimension of 5 × 0.45 mm by using a TLC scanner (CAMAG). The results were generated by using HPTLC software WinCATS 1.4.4.6337. The phytotransformed dye SRR was characterized by FTIR (Agilent Cary 630 FTIR spectrometer) and compared with the control sample. The FTIR analysis was conducted in the mid-IR region of 400–4000 cm−1. The samples were mixed with spectroscopically pure KBr in a ratio of 5:95, pellets were put on the sample analyzer, and the analyses were carried out.
:2). After development, the plate was observed in a UV chamber (CAMAG) and scanned at 254 nm with a slit dimension of 5 × 0.45 mm by using a TLC scanner (CAMAG). The results were generated by using HPTLC software WinCATS 1.4.4.6337. The phytotransformed dye SRR was characterized by FTIR (Agilent Cary 630 FTIR spectrometer) and compared with the control sample. The FTIR analysis was conducted in the mid-IR region of 400–4000 cm−1. The samples were mixed with spectroscopically pure KBr in a ratio of 5:95, pellets were put on the sample analyzer, and the analyses were carried out.
2.10. Phytotoxicity study
An SRR solution at 2000 ppm concentration in distilled water was prepared and applied for the toxicity testing on seeds of Sorghum vulgare, Phaseolus mungo and Vigna radiata at room temperature. Similarly, treated and untreated dye mixtures and effluents were used for phytotoxicity tests. The above mentioned three crop plants are commonly cultivated in India. 5 mL distilled water was used as a control. The seeds were placed in 5 mL solutions of dye and degraded metabolites separately at room temperature. The seed germination percentage was computed and the shoot length (plumule) and root length (radicle) were measured after 8 days.2.11. Statistical analysis
Data was analyzed by one-way analysis of variance (ANOVA) with Tukey–Kramer multiple comparisons test. Readings were considered significant when P was ≤0.05.3. Results and discussion
3.1. Decolorization of dyes by I. hederifolia L
The plants of I. hederifolia from a contaminated site showed different decolorization potentials for various dyes. I. hederifolia decolorized Navy Blue HE2R up to 96%, Green HE4B up to 90%, Blue 2RNL up to 89%, Direct Red 5B up to 87%, Reactive Red 2 up to 68%, Orange HE2R up to 56% and SRR up to 96% within 60 h. I. hederifolia showed versatility in decolorization of the dyes and showed the potential to be used for textile dyes treatment. Tissue cultures of I. hederifolia also afforded 90% decolorization of SRR within 96 h showing the direct role of this plant without any bacterial involvement. A UV-Vis scan (800–400 nm) of medium supernatants withdrawn after 96 h of exposure of I. hederifolia cultures to the dye SRR indicated its decolorization and decrease in concentration (Fig. 1). The tissue culture is recognized as an efficient tool for phytoremediation studies (Doran et al., 2009).29 Phytoremediation of textile dyes has earlier been shown by tissue cultures of P. grandiflora, Z. angustifolia, Blumea malcolmii and Sesuvium portulacastrum.5,6,10,13,30 | ||
| Fig. 1 UV-Vis spectrophotometric analysis of Scarlet RR, root exudates of Ipomoea hederifolia and the metabolites of the dye after treatment. | ||
3.2. Characterization of dye mixture and effluent before and after treatment
Textile dyes containing effluents are known to have high BOD and COD. Simulated dye mixture and textile effluent were also decolorized efficiently by I. hederifolia, and it afforded ADMI removal of 85% and 88%, BOD removal of 65% and 63% and COD removal of 62% and 68%, respectively (Table 1). Effluents and mixtures of dyes do not have true colors, therefore, their color is measured by the tristimulus method, which measures the color value independent of the hues.12 It is highly desired to have a phytoremediator, which is able to treat actual textile effluents. Earlier, Phragmites australis, Aster amellus, Glandularia pulchella, Portulaca grandiflora, Petunia grandiflora, Zinnia angustifolia and Tagetes patula have been shown to treat the textile effluents and bring them down to harmless levels.11–15 I. hederifolia is a natural inhabitant of the dye effluent contaminated soils; therefore, it can be proposed for further on field phytoremediation trials.| Parameter | Untreated effluent | Treated effluent | Untreated dye mixture | Treated dye mixture |
|---|---|---|---|---|
| COD (mg L−1) | 1840 | 580 | 1430 | 540 |
| BOD (mg L−1) | 1630 | 590 | 1250 | 430 |
| ADMI units | 962 | 108 | 678 | 96 |
| Turbidity (NTU) | 209 | 56 | 87 | 35 |
| TDS (mg L−1) | 3478 | 998 | 94 | 36 |
| TSS (mg L−1) | 496 | 408 | 28 | 17 |
| Hardness | 410 | 130 | 380 | 100 |
| pH | 10.2 | 7.9 | 8.1 | 7.5 |
| Conductivity (S m−1) | 3.80 × 10−3 | 0.90 × 10−3 | 2.80 × 10−3 | 1.40 × 10−3 |
3.3. Anatomical studies of stems and roots during dye degradation
An anatomical study of stems for understanding the histochemistry, movement and metabolism of SRR was carried out. The control plant stem (Fig. 2a) showed no coloration of any cell and they appeared to be normal and undisturbed. After 6 h of SRR dye exposure (Fig. 2b), dye was found to be accumulated in the epidermal cells. As the exposure continued the accumulation increased, reaching up to the cortical cells at 12 h (Fig. 2c). Furthermore, at 24 h (Fig. 2d), the accumulation increased in the cortex along with disappearance of dye around epidermal layer, showing degradation and metabolism of SRR. Phytotransformation in epidermal, as well as cortical cells, after 48 h (Fig. 2e) clearly revealed increased dye removal. The exposed plants were transferred to normal tap water and the stem section regained the clear cortex with some distortion in epidermal cells having a small amount of dye in them (Fig. 2f). | ||
| Fig. 2 Anatomy of stem of I. hederifolia (a) control plant, SRR exposed plants (b) at 6 h, (c) 12 h, (d) 24 h, (e) 48 h and (f) plants exposed to normal tap water after 48 h of SRR exposure. | ||
The meristematic cells of I. hederifolia were observed under a microscope, and the control plant root tip cells (Fig. 3a) showed normal cuboidal cells but after dye exposure the cells were observed to be elongated in shape (Fig. 3b). This distortion in morphology could have occurred because of the dye stress.
 | ||
| Fig. 3 Meristematic cells of root tips of I. hederifolia (a) control plant (unexposed to Scarlet RR), (b) plant exposed to Scarlet RR. | ||
3.4. Photosynthetic pigments
Photosynthetic pigments are considered to be one of the factors sensitive to stress and the energy requirement of cells. After 60 h, the chlorophyll (a, b) and carotenoid concentrations were estimated. Surprisingly, the chlorophyll and carotenoid concentrations were found to be increased in the plants exposed to SRR compared to those in control plants, which were kept in normal tap water (Table 3). I. hederifolia is a C4 plant, which under abiotic stress like the presence of dye might have synthesized more chlorophylls and carotenoids, leading to an increase in their concentration in the exposed plants. Similar kinds of results were reported previously where temperature played a role as an abiotic stress in which a decrease in photosynthetic pigments were seen in the case of C3 plants and an increase in pigment concentration was observed in the case of C4 plants.31| Enzymes | I. hederifolia root cells | I. hederifolia shoot cells | ||
|---|---|---|---|---|
| Control | Test | Control | Test | |
| a Values are a mean of three experiments ± SEM. Significantly different from control (0 h) at †P < 0.05, ††P < 0.01 and †††P < 0.001 by one-way ANOVA with Tukey–Kramer comparison test.b Activity in units min−1 mg−1.c μg of DCIP reduced min−1 mg protein−1.d μg of Azo dye reduced min−1 mg protein−1.e 50% inhibition of the NBT photoreduction rate (U mg−1 protein).f Nano moles of H2O2 Utilized (U mg−1 protein).g NA: no activity. | ||||
| Lignin peroxidaseb | 0.25 ± 0.01 | 1.82 ± 0.04††† | 0.23 ± 0.65 | 0.70 ± 0.11†† |
| Laccaseb | 13.86 ± 0.35 | 18.14 ± 0.28† | 0.29 ± 0.95 | 0.91 ± 0.87† |
| DCIP reductasec | 256.02 ± 30.41 | 407.6 ± 1.45††† | 350.55 ± 0.35 | 495.44 ± 0.16†† |
| Tyrosinaseb | 40.24 ± 0.87 | 49.46 ± 0.53† | 16.28 ± 0.33 | 46.14 ± 0.26††† |
| Azo reductased | NAg | NAg | 16.39 ± 0.057 | 19.27 ± 0.41† |
| Veratryl alcohol oxidaseb | 1.47 ± 0.13 | 3.62 ± 0.13†† | 3.66 ± 0.88 | 4.89 ± 0.16† |
| Catalasee | 0.046 ± 0.76 | 0.081 ± 0.56† | 0.35 ± 0.49 | 1.3 ± 0.90†† |
| Superoxide dismutasef | 2.15 ± 0.38 | 3.81 ± 0.84†† | 1.27 ± 0.66 | 3.92 ± 0.58†† |
| Sample | Chlorophyll a (μg mL−1) | Chlorophyll b (μg mL−1) | Total chlorophyll content (μg mL−1) | Carotenoids (μg mL−1) |
|---|---|---|---|---|
| Control plant | 10.979 ± 0.68 | 7.162 ± 0.89 | 18.141 ± 0.76 | 13.412 ± 0.66 |
| Test plant | 17.952 ± 0.87 | 11.755 ± 1.12 | 29.707 ± 1.06 | 18.262 ± 0.90 |
3.5. Enzyme induction studies during textile dye degradation process
Enzymatic treatments possess no risk of biological contamination and do not have any harmful impact on the ecosystems. In order to have additional insight into the decolorization mechanism, the activities of lignin peroxidase, veratryl alcohol oxidase, tyrosinase, laccase, 2,6-dichlorophenol indophenol reductase and azo reductase were assayed. Some organic compounds can be precisely degraded and completely mineralized by plant and microbial enzymes.32 In this study, the root cells of in vitro grown I. hederifolia gave significant inductions in the enzyme activities of LiP (628%), veratryl alcohol oxidase (146%), DCIP reductase (59%), laccase (31%), tyrosinase (23%), catalase (76%) and SOD (77%). While in the case of stem cells, significant inductions in the enzyme activities of LiP (204%), veratryl alcohol oxidase (33%), DCIP reductase (41%), laccase (214%), tyrosinase (183%), catalase (271%) and SOD (209%) were seen (Table 2). Earlier, I. palmata and Saccharum spontaneum and Phragmites australis were used for dye decolorization, showing the involvement of peroxidase enzymes.33,34 B. malcolmii, A. amellus, Zinnia angustifolia, P. grandiflora and G. pulchella also showed an active role of veratryl alcohol oxidase, tyrosinase, LiP, laccase and DCIP reductase during the decolorization of Direct Red 5B, Remazol Red, Brilliant Blue R and Green HE4B.6,12–15,21 Induction in the activities of lignin peroxidase, laccase, NADH-DCIP reductase, veratryl alcohol oxidase and tyrosinase shows their direct or indirect role in degradation of SRR, showing their role in degradation.3.6. Analysis of metabolites
The FTIR spectrum of an untreated sample of SRR (Fig. 4a) showed the following peaks: peaks at 3094.8 and 2904.8 cm−1 showed C–H stretching, a peak at 3517.3 cm−1 represented O–H stretching, a peak at 2454.9 cm−1 showed NH+ stretching, a peak at 2247.6 cm−1 revealed C![[triple bond, length as m-dash]](https://www.rsc.org/images/entities/char_e002.gif) N stretching, a peak at 1598.5 cm−1 showed NO2 stretching, and at 1517.0 cm−1 represented N
N stretching, a peak at 1598.5 cm−1 showed NO2 stretching, and at 1517.0 cm−1 represented N![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O as in aromatic nitro compounds. The peak at 1043.1 cm−1 showed the sulfonated nature of SRR, the peaks at 808.7, 969.74 and 682.82 cm−1 showed C–H deformation, whereas 620.62 cm−1 showed C–Cl stretching. The treated sample of SRR (Fig. 4b) showed different peaks: 2947.8, 2835.1, 2798.5 cm−1 showed C–H stretching and 1115.6, 1021.1 cm−1 indicated the presence of C–OH stretching. Similarly, the untreated mixture of dyes (Fig. 4c) showed peaks at 3425.7 and 2327.6 cm−1, revealing N–H stretching, a peak at 1497.7 cm−1, which showed CC and CN stretching, a peak at 1544.2 cm−1, which showed NO stretching, peaks at 1408 and 1109.4 cm−1 stood for C–H deformation, a peak at 1371.4 cm−1 revealed SO stretching, a peak at 1324.9 cm−1 showed NO2 stretching, peaks at 1181.8 and 909.7 cm−1 showed C–OH stretching, a peak at 735.3 cm−1 represented the aromaticity of the compounds, whereas a single peak at 664.8 cm−1 showed C–Cl stretching. The treated sample of dye mixture (Fig. 4d) showed a peak at 3233.2 cm−1, representing NO stretching, a peak at 2957.6 cm−1, showing C–H stretching, peaks at 1445.2, 1303.5 and 918.2 cm−1, which showed C–H deformation, a peak at 1227.2 cm−1, which showed SO stretching, and a peak at 695.2 cm−1, which showed C–Cl stretching. The FTIR spectrum of the untreated effluent sample (Fig. 4e) showed a peak at 3442.8 cm−1 for N–H stretching, peaks at 2927.1 cm−1 and 2850.8 cm−1 for C–H stretching, a peak at 1635.6 cm−1, showing CC stretching, a peak at 1591.6 cm−1, showing N–H deformation, a peak at 1495.3 cm−1, showing NO stretching, peaks at 1340.4 and 1132 cm−1, showed SO stretching, peaks at 1221.9, 844.5 and 735.6 cm−1, which represented C–H deformation, a peak at 1015.5 cm−1, which showed C–OH stretching and a peak at 637.9 cm−1 revealed the C–Cl stretching. On the other hand, the FTIR spectrum of the treated effluent sample (Fig. 4f) showed different peaks at 3417.8 cm−1, showing N–H stretching, the peaks at 2922.7 and 2852.1 cm−1, which showed C–H stretching, a peak at 1613.7 cm−1, which showed CO stretching, and a peak at 1420.6 cm−1, which showed SO stretching. The differential FTIR spectra obtained before and after the treatment of SSR, dye mixture and real textile effluent supported the transformation of the dyes to different products. This also reveals the phytotransformation potential of I. hederifolia.
O as in aromatic nitro compounds. The peak at 1043.1 cm−1 showed the sulfonated nature of SRR, the peaks at 808.7, 969.74 and 682.82 cm−1 showed C–H deformation, whereas 620.62 cm−1 showed C–Cl stretching. The treated sample of SRR (Fig. 4b) showed different peaks: 2947.8, 2835.1, 2798.5 cm−1 showed C–H stretching and 1115.6, 1021.1 cm−1 indicated the presence of C–OH stretching. Similarly, the untreated mixture of dyes (Fig. 4c) showed peaks at 3425.7 and 2327.6 cm−1, revealing N–H stretching, a peak at 1497.7 cm−1, which showed CC and CN stretching, a peak at 1544.2 cm−1, which showed NO stretching, peaks at 1408 and 1109.4 cm−1 stood for C–H deformation, a peak at 1371.4 cm−1 revealed SO stretching, a peak at 1324.9 cm−1 showed NO2 stretching, peaks at 1181.8 and 909.7 cm−1 showed C–OH stretching, a peak at 735.3 cm−1 represented the aromaticity of the compounds, whereas a single peak at 664.8 cm−1 showed C–Cl stretching. The treated sample of dye mixture (Fig. 4d) showed a peak at 3233.2 cm−1, representing NO stretching, a peak at 2957.6 cm−1, showing C–H stretching, peaks at 1445.2, 1303.5 and 918.2 cm−1, which showed C–H deformation, a peak at 1227.2 cm−1, which showed SO stretching, and a peak at 695.2 cm−1, which showed C–Cl stretching. The FTIR spectrum of the untreated effluent sample (Fig. 4e) showed a peak at 3442.8 cm−1 for N–H stretching, peaks at 2927.1 cm−1 and 2850.8 cm−1 for C–H stretching, a peak at 1635.6 cm−1, showing CC stretching, a peak at 1591.6 cm−1, showing N–H deformation, a peak at 1495.3 cm−1, showing NO stretching, peaks at 1340.4 and 1132 cm−1, showed SO stretching, peaks at 1221.9, 844.5 and 735.6 cm−1, which represented C–H deformation, a peak at 1015.5 cm−1, which showed C–OH stretching and a peak at 637.9 cm−1 revealed the C–Cl stretching. On the other hand, the FTIR spectrum of the treated effluent sample (Fig. 4f) showed different peaks at 3417.8 cm−1, showing N–H stretching, the peaks at 2922.7 and 2852.1 cm−1, which showed C–H stretching, a peak at 1613.7 cm−1, which showed CO stretching, and a peak at 1420.6 cm−1, which showed SO stretching. The differential FTIR spectra obtained before and after the treatment of SSR, dye mixture and real textile effluent supported the transformation of the dyes to different products. This also reveals the phytotransformation potential of I. hederifolia.
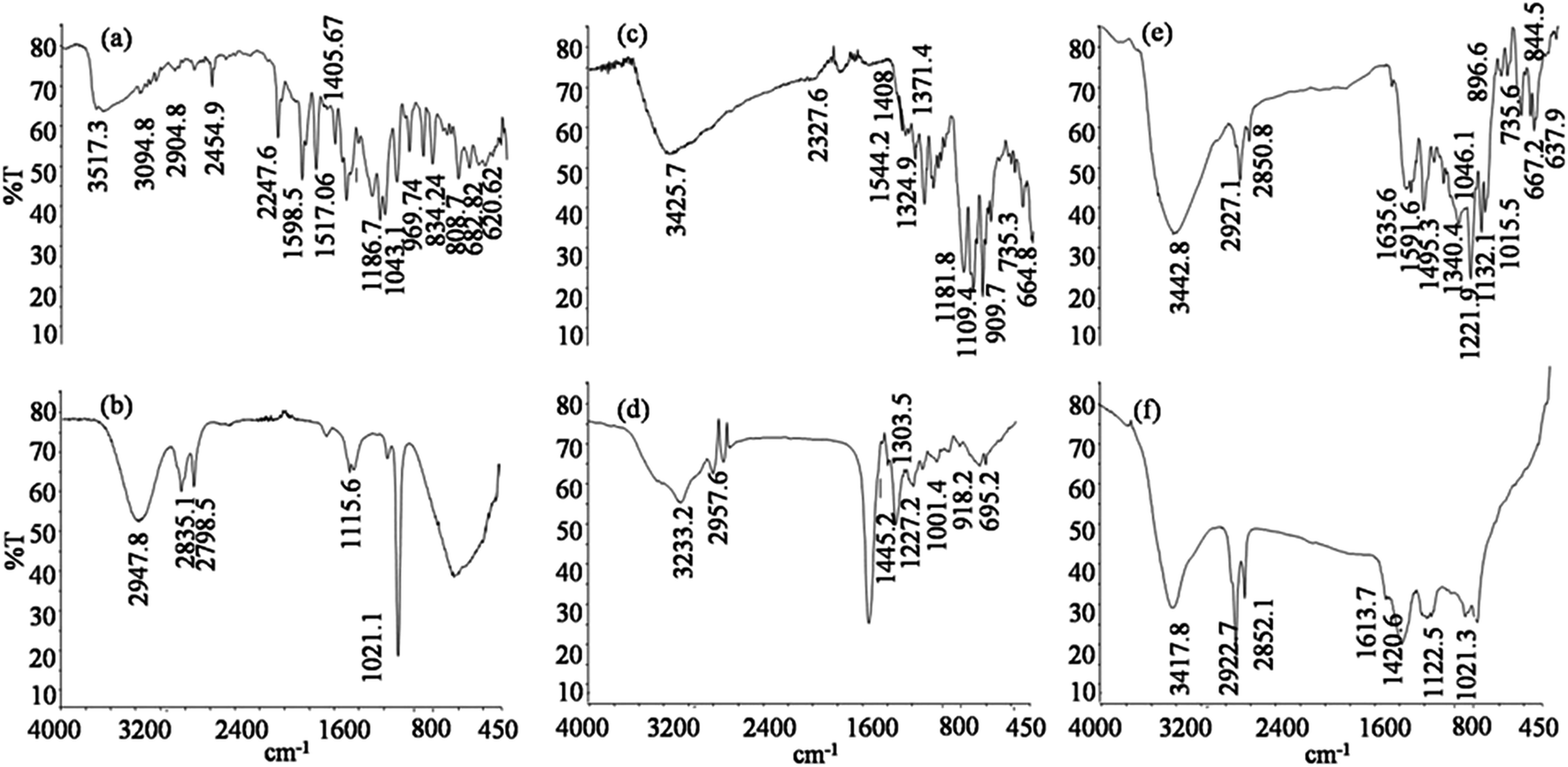 | ||
| Fig. 4 FTIR analysis of (a) Scarlet RR, (b) metabolites of Scarlet RR, (c) untreated dye mixture, (d) treated dye mixture, (e) untreated effluent and (f) effluent after treatment. | ||
HPLC analysis of the untreated SRR (Fig. 5a) showed the peak at a retention time of 3.12 min, whereas this peak disappeared after treatment (Fig. 5b) with the appearance of new peaks at retention times of 2.10, 2.51, 3.03, 3.31, 3.58 and 4.50 min. HPLC analysis of the mixture of dyes (Fig. 5c) showed the peaks at retention times of 1.90, 2.43, 2.59, 2.90, 3.28 and 5.69 min, while the treated dye mixture (Fig. 5d) showed peaks at retention times of 2.00, 2.11, 2.57, 3.33, 3.80 and 4.50 min. Similarly, the HPLC spectrum of untreated effluent (Fig. 5e) showed distinct peaks at retention times of 3.05, 3.39, 3.68, 4.28, 4.55 and 6.32 min. However, treated effluent (Fig. 5f) showed peaks at retention times of 3.14, 3.52 and 3.62 min. The differential HPLC spectra obtained in untreated and treated samples of SRR, the mixture of dyes and the textile effluent confirms the photodegradation of dyes.
 | ||
| Fig. 5 HPLC analysis (a) Scarlet RR, (b) metabolites of Scarlet RR, (c) untreated dye mixture, (d) treated dye mixture, (e) untreated effluent and (f) effluent after treatment. | ||
HPTLC analysis (Fig. 6) of the untreated SRR (Lane 1) showed three peaks at an Rf value of 0.85 with a major band at 0.76 and a third band at 0.53 with an absorbance of 0.4, 0.2 and 0.9 AU. In lane 2 (treated SRR) only a diminished peak was observed at 0.85 Rf with an absorbance of 0.2 AU, confirming the biodegradation of SRR. Lane 3 (untreated dye mixture) showed a distinguished appearance of 5 bands at Rf values of 0.87, 0.67, 0.38, 0.20 and 0.16 with absorbances of 138.4, 95.5, 29.5, 13.9 and 97.6 AU. Lane 4 (treated dye mixture) showed a mere presence of dye at Rf values of 0.67 and 0.2 having absorbances of 23.1 and 16.8 AU. Untreated textile effluent (Lane 5) showed 5 bands at Rf values of 0.54, 0.41, 0.29, 0.21 and 0.12 with absorbances of 29.8, 76.4, 494.2, 279.0 and 14.7 AU, whereas treated effluent showed removal of most of the peaks and only three peaks were seen at Rf values of 0.96, 0.42 and 0.30 with absorbances of 69.5, 18.3 and 82.7 AU.
 | ||
| Fig. 6 HPTLC plate image with 3-D spectral scan at 254 nm. | ||
GCMS analysis was performed to know the extracted metabolites of SRR. Six separate peaks were obtained. The pathway of degradation of SRR was predicted with the help of metabolites obtained and the analysis of induction in the activities of the enzymes in the roots of I. hederifolia Linn. SRR undergoes asymmetric cleavage by the action of LiP to form two intermediate compounds, quinolin-4(3H)-one and 2-(methylamino)-N-(4-nitro-1l3-chlorinin-2-yl) acetamide. The ring of quinolin-4(3H)-one undergoes cleavage by the action of tyrosinase to form 2-ethenyl-2,5-dihydropyridin-4(3H)-one, which further undergoes reduction and de-ethenylation to afford 2,3,4,5-tetrahydropyridine. 2-(methylamino)-N-(4-nitro-1l3-chlorinin-2-yl)acetamide undergoes oxidative cleavage by laccase to afford 2-amino-N-(4-hydroxy-1l3-chlorinin-2-yl) acetamide, which further upon deamination and reduction gives N-methyl-1l3-chlorinin-2-amine (Fig. 7). Lignin peroxidase and veratryl alcohol oxidase are known to cleave the dye molecules asymmetrically.12–16,30 The laccase cleaves the dyes by oxidative cleavage.35 The differential enzymatic status also influences the degradation mechanism. At natural sites of dye contaminated soils, other factors such as soil microflora and environmental conditions along with plants may act on the dye molecule synergistically and mineralization of the dye molecules may occur. The I. hederifolia showed great potential to degrade SRR dye to different metabolites.
 | ||
| Fig. 7 Proposed pathway for degradation of Scarlet RR by Ipomoea hederifolia L. | ||
3.7. Phytotoxicity analysis
The textile dye SRR, mixture of dyes and effluent, showed inhibition of germination of Phaeseolus mungo, Sorghum vulgare and Vigna radiata seeds. The untreated textile dye SRR, mixture of dyes and effluent, showed 60%, 50% and 60% inhibition of P. mungo seed germination respectively, whereas S. vulgare seeds showed 60%, 50%, 60% inhibition of germination, and for V. radiata germination inhibition percentage was found to be 50%, 60% and 60%, respectively. The plumule and radicle lengths also decreased in the case of untreated textile dye SRR, effluent and dye mixture samples. The eight days of exposure of the untreated textile dye SRR, textile effluent and dye mixture were highly toxic to the plantlets as the root and shoot lengths were also inhibited when compared to treated effluent and dye mixture samples. The treated samples were found to be less toxic and showed lower germination inhibition percentages when compared to their untreated forms. A significant increase in seed germination percentage and, shoot and root lengths reveals the reduced toxicity of the metabolites formed after the treatment by I. hederifolia (Table 4).| Parameters studied | Water | Scarlet RR | Scarlet RR metabolites | Untreated dye mixture | Treated dye mixture | Untreated textile effluent | Treated textile effluent |
|---|---|---|---|---|---|---|---|
| a Values are a mean of three experiments ± SE. Root and shoot lengths of plants grown in textile dye SRR, textile effluent and mixture of dyes, respectively, are significantly different from that of plants grown in distilled water by †P < 0.001. Root and shoot lengths of plants grown in the degraded treated SRR, textile effluent and mixture of dyes, respectively, are also significantly different from that of plants grown in untreated SRR, textile effluent and mixture of dyes by ‡P < 0.001. | |||||||
| Phaseolus mungo | |||||||
| Germination (%) | 100 | 40 | 100 | 50 | 80 | 40 | 70 |
| Plumule (cm) | 9.71 ± 0.05 | 3.53 ± 0.06† | 8.06 ± 0.03‡ | 2.83 ± 0.06† | 3.53 ± 0.03 ‡ | 2.62 ± 0.02† | 3.24 ± 0.04 ‡ |
| Radicle (cm) | 6.23 ± 0.03 | 2.42 ± 0.05† | 3.60 ± 0.05 ‡ | 3.2 ± 0.05† | 3.63 ± 0.02 ‡ | 2.86 ± 0.03† | 3.15 ± 0.02 ‡ |
| Sorghum vulgare | |||||||
| Germination (%) | 100 | 40 | 90 | 50 | 90 | 40 | 70 |
| Plumule (cm) | 7.26 ± 0.08 | 4.13 ± 0.06† | 5.81 ± 0.10 ‡ | 3.63 ± 0.08† | 4.3 ± 0.05 ‡ | 3.5 ± 0.06† | 4.33 ± 0.15 ‡ |
| Radicle (cm) | 5.94 ± 0.12 | 1.46 ± 0.04† | 2.80 ± 0.03 ‡ | 2.3 ± 0.05† | 2.8 ± 0.05 ‡ | 1.8 ± 0.01† | 2.41 ± 0.02 ‡ |
| Vigna radiata | |||||||
| Germination (%) | 90 | 50 | 80 | 40 | 80 | 40 | 80 |
| Plumule (cm) | 5.93 ± 0.08 | 2.73 ± 0.06† | 5.83 ± 0.06 ‡ | 2.53 ± 0.03† | 5.73 ± 0.03 ‡ | 2.36 ± 0.08† | 5.83 ± 0.06 ‡ |
| Radicle (cm) | 2.46 ± 0.06 | 1.02 ± 0.90† | 2.36 ± 0.02 ‡ | 0.93 ± 0.90† | 2.3 ± 0.06 ‡ | 0.73 ± 0.06† | 1.13 ± 0.03 ‡ |
4. Conclusion
The phytoremediation of textile dyes has mainly been carried out by using common garden plants whose potential is yet to be tested for the in situ remediation purpose. This study explores the phytoremediation potential of I. hederifolia, a plant inhabitant of the actual dye disposal site. Its seeds germinate normally in high dye concentration and can complete the life cycle in high dye load. The plant could decolorize a real textile effluent sample and a dye mixture to meaningful levels. This study clearly revealed the dye removal by I. hederifolia at anatomical level. Studies to explore this plant in a constructed wetland system for textile effluent treatment at an industrial scale are underway.Acknowledgements
Niraj R. Rane, Avinash G. Gholave, Shrirang R. Yadav and Sanjay P. Govindwar wish to thank Department of Biotechnology (DBT), New Delhi, India, for the funding under Interdisciplinary Program for Life Science (IPLS) project to Shivaji University, Kolhapur.References
- A. Khataee, G. Dehghan, A. Ebadi, M. Zarei and M. Pourhassan, Bioresour. Technol., 2010, 101, 2252–2258 CrossRef CAS PubMed.
- A. Kabra, R. Khandare and S. Govindwar, Water Res., 2013, 47, 1035–1048 CrossRef CAS PubMed.
- R. Khandare, A. Kabra, A. Kadam and S. Govindwar, Int. Biodeterior. Biodegrad., 2013a, 78, 89–97 Search PubMed.
- T. Bhalerao and P. Puranik, Int. Biodeterior. Biodegrad., 2007, 59, 315–321 CrossRef CAS.
- R. Khandare, A. Kabra, A. Awate and S. Govindwar, Int. J. Environ. Sci. Technol., 2013b, 10, 1039–1050 Search PubMed.
- A. Kagalkar, U. Jagtap, J. Jadhav, V. Bapat and S. Govindwar, Bioresour. Technol., 2009, 100, 4104–4110 CrossRef CAS PubMed.
- A. Dietz and J. Schnoor, Environ. Health Perspect., 2001, 109, 163–168 CrossRef CAS PubMed.
- S. Aubert and J. Schwitzguébel, Water Res., 2004, 38, 3569–3575 CrossRef CAS PubMed.
- L. Davies, C. Carias, J. Novais and S. Martins-Dias, Ecol. Eng., 2005, 25, 594–605 CrossRef.
- A. Kagalkar, M. Jadhav, V. Bapat and S. Govindwar, Bioresour. Technol., 2011, 102, 10312–10318 CrossRef CAS PubMed.
- S. Ong, K. Uchiyamam, D. Inadama, Y. Ishida and K. Yamagiwa, Bioresour. Technol., 2011, 101, 9049–9057 CrossRef PubMed.
- R. Khandare, A. Kabra, D. Tamboli and S. Govindwar, Chemosphere, 2011a, 82, 1147–1154 Search PubMed.
- R. Khandare, A. Kabra, M. Kurade and S. Govindwar, Bioresour. Technol., 2011b, 102, 6774–6777 Search PubMed.
- A. Kabra, R. Khandare, M. Kurade and S. Govindwar, Environ. Sci. Pollut. Res., 2011a, 18, 1360–1373 Search PubMed.
- A. Kabra, R. Khandare, T. Waghmode and S. Govindwar, J. Hazard. Mater., 2011b, 190, 424–431 Search PubMed.
- A. Patil and J. Jadhav, Chemosphere, 2013, 92, 225–232 CrossRef CAS.
- A. Watharkar, R. Khandare, A. Kamble, A. Mulla, S. Govindwar and J. Jadhav, Environ. Sci. Pollut. Res., 2013a, 20, 939–949 Search PubMed.
- A. Watharkar, N. Rane, S. Patil, R. Khandare and J. Jadhav, Bioresour. Technol., 2013, 142, 246–254 CrossRef CAS.
- American Public Health Association (APHA), APHA–AWWA–WEF, Washington, DC, USA, 20th edn, 1998.
- D. Arnon, Plant Physiol., 1949, 24, 1–15 CrossRef CAS PubMed.
- R. Khandare, N. Rane, T. Waghmode and S. Govindwar, Environ. Sci. Pollut. Res., 2012, 19, 1709–1718 CrossRef CAS PubMed.
- V. Shanmugam, M. Kumari and K. Yadav, Indian J. Biochem. Biophys., 1999, 36, 39–43 CAS.
- N. Hatvani and I. Mecs, Process Biochem., 2001, 37, 491–496 CrossRef.
- X. Zhang and W. Flurkey, J. Food Sci., 1997, 62, 97–100 CrossRef CAS.
- M. Salokhe and S. Govindwar, World J. Microbiol. Biotechnol., 1999, 1, 229–232 Search PubMed.
- R. Bourbonnais and M. Paice, Biochem. J., 1988, 255, 445–450 CrossRef CAS.
- B. Acharya, S. Mohanna and D. Madamwar, Bioresour. Technol., 2007, 99, 4621–4626 CrossRef PubMed.
- O. Lowry, N. Rosebrough, A. Farr and R. Randall, J. Biol. Chem., 1951, 193, 265–275 CAS.
- P. Doran, Biotechnol. Bioeng., 2009, 103, 60–76 CrossRef CAS PubMed.
- A. Patil, V. Lokhande, P. Suprasanna, V. Bapat and J. Jadhav, Planta, 2012, 235, 1051–1063 CrossRef CAS PubMed.
- A. Taylor and J. Rowley, Plant Physiol., 1971, 47, 713–718 CrossRef CAS.
- E. Wild, G. Thomas and K. Jones, Environ. Sci. Technol., 2005, 39, 3695–3702 CrossRef CAS PubMed.
- T. Shaffiqu, J. Roy, R. Nair and T. Abraham, Appl. Biochem. Biotechnol., 2002, 103, 315–326 CrossRef.
- C. Carias, J. Novais and S. Martins-Dias, Water Sci. Technol., 2007, 56, 263–269 CrossRef CAS PubMed.
- M. Chivukula and V. Renganathan, Appl. Environ. Microbiol., 1995, 61, 4374–4377 CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra06840h |
| This journal is © The Royal Society of Chemistry 2014 |