
Elucidation of the chemical composition of avian melanin†
Abstract
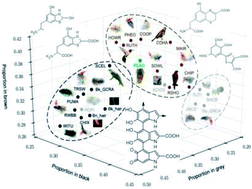
Our understanding of the chemical composition of melanin remains limited, due to a paucity of direct measurements. Avian feathers have an unparalleled diversity of melanin-based color mirroring their complex chemistry. Synchrotron-based photoionization mass spectrometry is used to determine the chemical composition of melanin from samples of black, brown, grey and iridescent feathers.
 Please wait while we load your content...
Please wait while we load your content...

