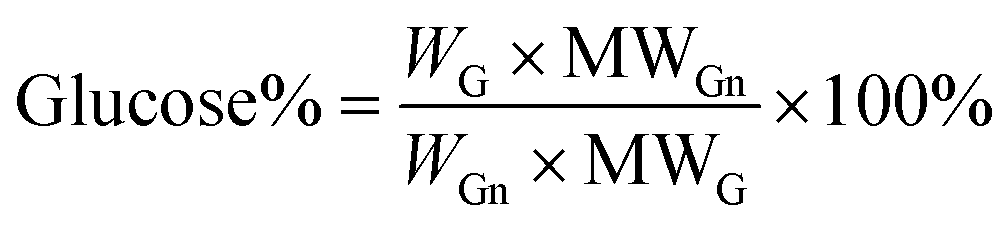DOI:
10.1039/C4RA01631A
(Paper)
RSC Adv., 2014,
4, 23492-23504
A comprehensive mechanistic kinetic model for dilute acid hydrolysis of switchgrass cellulose to glucose, 5-HMF and levulinic acid
Received
25th February 2014
, Accepted 16th May 2014
First published on 16th May 2014
Abstract
Switchgrass was treated by 1% (w/w) H2SO4 in batch tube reactors at temperatures ranging from 140–220 °C for up to 60 minutes. In this study, release patterns of glucose, 5-hydroxymethylfurfural (5-HMF), and levulinic acid from switchgrass cellulose were investigated through a mechanistic kinetic model. The predictions were consistent with the measured products of interest when new parameters reflecting the effects of reaction limitations, such as cellulose crystallinity, acid soluble lignin–glucose complex (ASL–glucose) and humins that cannot be quantitatively analyzed, were included. The new mechanistic kinetic model incorporating these parameters simulated the experimental data with R2 above 0.97. Results showed that glucose yield was most sensitive to variations in the parameter regarding the cellulose crystallinity at low temperatures (140–180 °C), while the impact of crystallinity on the glucose yield became imperceptible at elevated temperatures (200–220 °C). Parameters related to the undesired products (e.g. ASL–glucose and humins) were the most sensitive factors compared with rate constants and other additional parameters in impacting the levulinic acid yield at elevated temperatures (200–220 °C), while their impacts were negligible at 140–180 °C. These new findings provide a more rational explanation for the kinetic changes in dilute acid pretreatment performance and suggest that the influences of cellulose crystallinity and undesired products including ASL–glucose and humins play key roles in determining the generation of glucose, 5-HMF and levulinic acid from biomass-derived cellulose.
1. Introduction
Pretreatment has been the subject of considerable R&D, and a host of biological, chemical, and physical methods have been researched. Overall, most of this past effort has been directed at maximizing sugar yields from hemicellulose and cellulose through trials with different chemicals, operating conditions, and equipment configurations. Technology has recently been developed to catalytically convert biomass sugars and their sequential degradation products (e.g. 5-HMF, levulinic acid, furfural and formic acid) into “drop-in” fuel components, including gasoline and jet and diesel fuels, which are compatible with conventional fuels.1 However, pretreatment is still considered to be one of the most costly steps in lignocelluloses conversion process. The most cost effective pretreatment to date is dilute acid pretreatment because of its ability to achieve high yields of reactive intermediates from both cellulose and hemicelluloses.2–4
Cellulose, as the major polymer from biomass, is the potential source for many kinds of chemicals such as glucose, 5-HMF, and levulinic acid etc.3 Dilute acid hydrolysis of lignocellulosic biomass is a widely used technology for generating these cellulose-derived compounds.3,5 However, the yields of glucose, 5-HMF and levulinic acid obtained from pretreatment in previous studies were limited to some extent. The glucose yields from dilute acid hydrolysis of lignocellulosic biomass reported in previous research were often limited to 65–70%.6 Alternatively, employing a simulated countercurrent shrinking bed system led to a higher than 85% of glucose yield.7 5-HMF, which is unstable under acidic conditions and can be further rehydrated into levulinic acid, was the sequential degradation product via dehydration of glucose.3 It was reported that less than 10% 5-HMF was obtained in dilute acid (e.g. 3.3% (w/w) hydrochloric acid) treatments of cellulose at temperatures ranging from 160–200 °C.8 Levulinic acid is much more stable than its precursor 5-HMF under acidic conditions.3 However, levulinic acid yields reported in previous studies did not exceed 60%.3,8,9 Previous kinetic studies regarding the generation of glucose, 5-HMF and levulinic acid in dilute acidic conditions implied that the limited yields of these cellulosic derived compounds can be interpreted by the ratio of the rate constants regarding the products formation and degradation based on a simplified cellulose degradation pathway:3,5,10
However, apart from the kinetic interactions among various reaction steps based on this simplified cellulose degradation pathway, the limited yields of these derived products could be attributed to many other factors. Xiang et al.11 revealed that cellulose hydrolysis occurred slowly compared to amorphous polysaccharides (e.g. starch) under dilute acid conditions (120 °C, 4% (w/w) H2SO4). The crystallinity of cellulose played a significant role in resisting the dilute acid hydrolysis of cellulose.12 Furthermore, the formation of undesired byproducts during cellulose decomposition also reduced the yields of glucose, 5-HMF and levulinic acid. It was reported that acid soluble lignin (ASL) can be generated during dilute acid pretreatment of lignocellulosic biomass.13 Glucose was prone to react with ASL through a recombination reaction in the acidic medium to form ASL–glucose.13 The formation of ASL–glucose not only decreases the glucose yield, but also is likely to reduce the production of glucose degradation products, i.e. 5-HMF and levulinic acid. The acid-catalyzed decomposition reactions of cellulose also produced an insoluble-solid product known as humins,8,9 which was observed by Patil and Lund14 as the degradation product of 5-HMF under 135 °C in water phase. Based on this, the yields of both 5-HMF and its subsequent product levulinic acid could be limited by the formation of humins under acidic conditions.9
Previous studies implied that the crystallinity of cellulose, as well as the formation of undesired products like ASL–glucose and humins are resistant to the acid hydrolysis15 or consume the production of glucose, 5-HMF and levulinic acid,13,14 thereby resulting in the reduced yields of products of interest. In this regard, investigating the effects of these potential limiting factors (e.g. crystallinity of cellulose, ASL–glucose and humins) on the production of glucose, 5-HMF and levulinic acid from cellulose could provide new insights into interactions among the various reaction steps that favor the formation of these cellulose-derived compounds. The crystallinity index (CrI) of cellulose can be observed by multiple methods such as X-ray diffraction technology.16 However, determining the CrI of cellulose from lignocellulosic biomass is not practical. Because the other two components in lignocellulosic biomass (i.e. hemicellulose and lignin) lack any regular crystal structure, the interference of these two compounds could result in the decrease of observed CrI of biomass derived cellulose compared with the pure cellulose.17 Although the qualitative analysis of some undesired byproducts (e.g. humins) has been reported through using advanced instruments such as scanning electron microscopy (SEM) and infrared (IR) spectra, the quantitative analytical approach of these reaction limitations (e.g. humins and ASL–glucose) was seldom reported in previous studies.13,14
Kinetic models of dilute acid pretreatment are vital to provide a foundation for understanding cellulose hydrolysis and the cause of enhanced performance by batch systems.18 Thus, developing mathematical interpretation of the cellulose degradation with the incorporation of parameters reflecting these reaction limitations could provide in-depth understanding about their influence on the generation of glucose, 5-HMF and levulinic acid etc. The current reaction models describing the acid-catalyzed cellulose hydrolysis to glucose were adapted from Saeman's first-order pseudo-homogeneous kinetic model of cellulose hydrolysis in a dilute acid batch system.19 It was a two-step consecutive first-order reaction: cellulose is first hydrolyzed into glucose, and then further converted into degradation products. Over the years, Saeman's model was applied by many researchers to study the acid-catalyzed hydrolysis of cellulose/lignocellulosic biomass under a wide range of reaction conditions, i.e. acid concentrations (0.05% (w/w)–8% (w/w)) and temperatures (90–240 °C), to predict the glucose yield generated from cellulose.13,20–22 A few modifications of such models have been made through adding new parameters in order to more precisely describe the mechanism of cellulose hydrolysis. Aguilar et al.23 proposed a two fraction model of cellulose hydrolysis (100–128 °C, 2% (w/w)–6% (w/w) H2SO4) based on the assumption that cellulose displayed a biphasic tendency with one portion of cellulose hydrolyzed fast and the other portion decomposed at a slower rate. This model is a modified Saeman's model with the introduction of a parameter reflecting the ratio of fast fraction to slow fraction. Orozco et al.24 applied this model to investigate the cellulose hydrolysis from grass clippings covering a wide range of reaction conditions: 135–200 °C with 0.4% (w/w)–10% (w/w) phosphoric acid. The ratio of fast fraction to slow fraction was found at the range of 0.5–0.8. Although the two-fraction model successfully fitted the experimental data (e.g. R2 = 0.97), limited direct evidence supported the existence of fast and slow portions in cellulose. These kinetic models proposed in previous studies mainly focused on describing the glucose generation from cellulose. Only a few kinetic models8,9 explained the complete acid-catalyzed cellulose degradation including the formation of 5-HMF and levulinic acid. Girisuta et al.9 and Shen and Wyman8 extended Saeman's model via incorporating 5-HMF and levulinic acid into the acid hydrolysis process of cellulose. The byproducts humins were included into the degradation pathway of cellulose in addition to 5-HMF and levulinic acid. The rate constants of varying reaction steps in determining the cellulose degradation process were compared and it revealed that the rate-controlling step shifted from levulinic acid formation initially to 5-HMF formation later. Nevertheless, the influence of undesired byproducts (e.g. humins) on the generation of glucose, 5-HMF and levulinic acid was still unknown although humins were symbolically introduced into the cellulose degradation pathway.
Although multiple models have been devised over the years,8,25–28 similar to the model developed by Shen and Wyman,8 they all evolve from the same first-order kinetic representation. None reported in the literature incorporated parameters reflecting the reaction limitations from biomass recalcitrance (e.g. crystallinity of cellulose) or undesired byproducts (e.g. ASL–glucose and humins) to adequately describe the changes in observed performance for the degradation pathway of lignocellulosic biomass derived polysaccharides (e.g. cellulose). Continuous efforts should be made to develop kinetic models that incorporate parameters reflecting the aforementioned crystallinity of cellulose, as well as undesired byproducts (e.g. ASL–glucose and humins) for describing the mechanism of cellulose degradation more precisely. In addition, the effects of these parameters on cellulose hydrolysis that are expected to be vital in determining the generation of value-added products glucose, 5-HMF and levulinic acid also need to be evaluated.
In this study, we conducted a kinetic study on the acid-catalyzed hydrolysis of switchgrass to cellulose-derived glucose, 5-HMF and levulinic acid with 1% (w/w) sulfuric acid under temperatures ranging from 140–220 °C for 0–60 min. A kinetic model integrating parameters representing the effects of crystallinity of cellulose, and the formation of undesired products (e.g. ASL–glucose, humin) was developed to provide a detailed description of cellulose acidic degradation mechanism as well as a successful simulation to the experimental data. The subsequent analysis of the sensitivity of the predicted glucose, 5-HMF and levulinic acid yields to changes in these parameter values provided new insights in the production of glucose, 5-HMF and levulinic acid from cellulose.
2. Kinetic models of switchgrass cellulose hydrolysis
2.1. Model I (conventional cellulose degradation model)
A conventional kinetic model8,9,19 with the incorporation of glucose, 5-HMF and levulinic acid reactions as shown in Scheme 1 was used to describe the cellulose hydrolysis process. This conventional model assumes irreversible pseudo-homogeneous first-order reactions and the rate equations can be presented as follows:| |
 | (1) |
| |
 | (2) |
| |
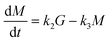 | (3) |
| |
 | (4) |
 |
| | Scheme 1 | |
The solutions of these rate expressions are given below:
| |
C = C0![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) exp(−k1t) exp(−k1t)
| (5) |
| |
 | (6) |
| |
 | (7) |
| |
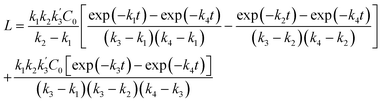 | (8) |
where
C,
G,
M and
L represent cellulose, glucose, 5-HMF and levulinic acid, which were expressed in the unit of yield (%).
26 C0 is the initial yield of cellulose (100%).
k1 (min
−1),
k2 (min
−1),
k3 (min
−1) and
k4 (min
−1) are the degradation rate constants of cellulose, glucose, 5-HMF and levulinic acid, respectively.
k′3 (min
−1) is the rate constant of levulinic acid formation.
2.2. Model II (modified cellulose degradation model)
A modified cellulose hydrolysis model was proposed as shown in Scheme 2. Three parameters α, β and γ were incorporated into this model to modify the conventional model:
 |
| | Scheme 2 | |
Parameter α represents the degree of cellulose crystallinity, which is expressed as the ratio of crystalline cellulose (Ccrystallinity) to total cellulose (C). The hydrolysis of cellulose is initiated by the cleavage of β-1,4-glycosidic bond of cellulose. However, the crystallinity of cellulose significantly hinders the hydronium ions from penetrating into cellulose and catalyzing the cleavage of the β-1,4-glycosidic bond.6,29,30 Consequently, only parts of the β-1,4-glycosidic bond in the cellulose linear structure possess the potential to be broken down. Introducing the parameter α into this kinetic model was intended to reflect the effects of the crystallinity on cellulose hydrolysis under varying reaction conditions. In this regard, the reactive portion of the cellulose that is unimpeded by crystallinity was defined as (1 − α) cellulose.
Parameter β represents the ratio of glucose transforming to the undesired products (e.g. ASL–glucose complex) (GASL) to overall generated glucose (G). Under acidic conditions, it was reported that additional pathways for glucose decomposition such as recombination reaction with ASL to form ASL–glucose existed.11,13 The presence of proton in acidic medium causes varying formations of reactive intermediates (e.g. protonized glucose), which would have high affinity for any positively charged molecules, including nucleophilic reaction partners (e.g. ASL).11,13,31 Thus, the active sites on ASL could lead to the condensation of ASL and glucose to form ASL–glucose. Based on this mechanism, with the introduction of parameter β, the portion of glucose compensated to undesired products (e.g. ASL–glucose) was therefore defined as (β) glucose, while the remainder was (1 − β) glucose. Parameter γ reflects the ratio of 5-HMF converted to byproducts (e.g. humins) (Mhumins) to overall generated 5-HMF (M). Previous studies reported that the 5-HMF acid-catalyzed conversion can be primarily described by two parallel reactions: one forming levulinic acid and the remainder forming humins.3,14 Patil and Lund14 demonstrated that the aldol addition and condensation with the involvement of 2,5-dioxo-6-hydroxy-hexanal as the intermediate are the primary reaction mechanism of the acidic catalyzed 5-HMF to humins. The formation of humins, which contain 55–65% carbon, 4–5% hydrogen, and 30–40% oxygen,32,33 contributed to the reduced levulinic acid yield. In this model, the portion of 5-HMF rehydrated to levulinic acid was defined as (1 − γ) 5-HMF, whereas those reacted to undesired byproducts (e.g. humins) was described as (γ) 5-HMF. Introducing these two parameters β and γ into the kinetic model allows the evaluation of the impact of the undesired byproducts (e.g. ASL–glucose and humins) on the production of 5-HMF and levulinic acid from lignocellulosic biomass.
The following assumptions are also made for the proposed model:
(1) The crystallized portion of cellulose is unreactive or reacts extremely slowly.34–38 Glucose and its subsequent degradation compounds are merely generated from the reactive portion of cellulose excluding the crystallized fractions.
(2) ASL–glucose is representative of the major undesired byproducts from glucose.11,13
(3) Humins represent the major undesired byproducts generated from 5-HMF.14
(4) Glucose, 5-HMF and levulinic acid are solely generated from cellulose. Other possible sources such as hemicellulose,39 starch40,41 and free sugars42,43 are excluded in this work since these compounds represent only a small source of glucose in switchgrass compared to cellulose.39–41,43
The kinetic model equations developed from Scheme 2 are described by eqn (9)–(12). Solutions to these equations have been obtained, providing explicit expressions for the yields of cellulose, glucose, 5-HMF and levulinic acid, as demonstrated by eqn (13)–(17).
| |
 | (9) |
| |
 | (10) |
| |
 | (11) |
| |
 | (12) |
| | |
C1−α = (1 − α)C0exp(−k1t)
| (13) |
| | |
C = Cα + C1−α = αC0 + (1 − α)C0exp(−k1t)
| (14) |
| |
 | (15) |
| |
 | (16) |
| |
 | (17) |
where
C,
G,
M and
L represent cellulose, glucose, 5-HMF and levulinic acid, which were expressed in the unit of yield (%).,
26 C0 is the initial yield of cellulose (100%).
Based on the aforementioned definition of α, β and γ, (1 − α), (1 − β) and (1 − γ) represent the ratio of reactive cellulose to cellulose, the ratio of glucose dehydrated to 5-HMF to overall generated glucose, and the ratio of 5-HMF rehydrated to levulinic acid to overall generated 5-HMF, respectively. In this regard, C(1 − α), (1 − β)G and (1 − γ)M representing the yield of reactive cellulose (%), the yield of glucose dehydrated to 5-HMF (%) and the yield of 5-HMF rehydrated to levulinic acid (%) are involved into the kinetic model eqn (9)–(12) instead of C, G and M to precisely depict the generation of glucose, 5-HMF and levulinic acid from cellulose.
3. Materials and methods
3.1. Feedstocks
Switchgrass was harvested in Prosser, Washington (July 2010, no. 13211). The composition of the switchgrass was determined based on Laboratory Analytical Procedure (LAP) of ‘‘Determination of Structural Carbohydrates and Lignin in Biomass’’.44 The switchgrass contains 40.2% glucan, 20.4% xylan, 2.8% arabinan, 1.1% galactan, 0.5% mannan, 19.5% Klason lignin and 6.6% ash on average basis. These materials were cut to shorter lengths, then grounded to a particle size of 200–400 μm with a laboratory mill (model: MF10 basic, IKA®works, Inc, NC, Wilmington) and then passed through a 40–60 mesh to obtain particles added in the reaction. All standard chemicals were purchased from Sigma-Aldrich, St. Louis, Mo.
3.2. Experimental methods
Batch tubular reactors (1.27 cm OD × 15.24 cm long with ×0.0889 cm wall thickness, Hastelloy C-276) (Swagelok Northwest, Richland, WA) were used in this study. The total volume of each reactor is 14.5 mL with 10 mL working volume. 0.5 g substrate was loaded into the batch tubular reactor with 10 mL of 1% (w/w) sulfuric acid. Reactors were heated to reach target temperature within 1 min in a 4 kW fluidized sand bath (model SBL-2D, Omega engineering, Inc., Stamford, CT) and maintained at temperature of 140–220 °C for 1–60 min. The reaction was quenched within 1 min by soaking the batch tubular reactor in cold water bath after being subjected to the target reaction temperature for a specified time. After the end, caps and plugs were removed, samples were pushed out and separated into liquid hydrolyzate and solid residue by vacuum filtration using a 0.22 μm glass fiber filter (Fisher Science, Pittsburgh, PA) for analysis.
3.3. Analytical methods
Glucose, 5-HMF, levulinic acid were analyzed through Dionex Ultimate 3000 HPLC equipped with a refractive detector (Shodex RI-101) and a ultimate 3000 autosampler using chromeleon 6.8 software (Dionex, Bannockburn, IL). Bio-Rad Aminex HPX-87H columns (Bio-Rad Laboratories, Hercules, CA) were operated under 65 °C. Mobile phase was 0.005 M H2SO4 with flow rate 0.6 mL min−1. Before analysis, the acid in the liquid hydrolyzate was neutralized with calcium carbonate until pH 5 to 6. The neutralized hydrolyzate was centrifuged and filtered for HPLC analysis.44 Yields of glucose, xylose, furfural, and 5-HMF were calculated as following:45| |
 | (18) |
| |
 | (19) |
| |
 | (20) |
In these equations, WGn is the initial weight of glucan (g per 100 g dw raw biomass), WG is the weight of glucose (g per 100 g dw raw biomass), W5-HMF is the weight of 5-HMF (g per 100 g dw raw biomass), WLA is the weight of levulinic acid (g per 100 g dw raw biomass). Molecular weight: MWGn = 162, MWG = 180, MW5-HMF = 126, MWLA = 116.
The cellulose in solid residue after separation was analyzed based on a standard analysis procedure developed by National Renewable Energy Laboratory (NREL),44 which is a two-step acid hydrolysis: the sample was treated with 72% (w/w) H2SO4 at 30 °C for 1 h; the reaction mixture was subsequently diluted to 4% (w/w) H2SO4 and autoclaved at 121 °C for 1 hour. The sugars in liquid after this two-step procedure were then determined by HPLC.
A MATLAB program was used to fit the parameters in eqn (5)–(8) and (13)–(17) to simulate the yields of cellulose residue, glucose, 5-HMF and levulinic acid obtained under tested conditions, respectively.
4. Results and discussion
4.1. Switchgrass cellulose hydrolysis
To study cellulose hydrolysis of switchgrass, we conducted a series of batch reactions under a wide temperature range of 140–220 °C within 0–60 min. Other conditions, including 1% (w/w) sulfuric acid, 40–60 mesh particle size of switchgrass and 5% (w/w) substrate concentration, were optimized in previous work.5,46 1% (w/w) sulfuric acid was selected because it was widely used for cellulose decomposition.5,47 Previous researches also revealed that particle size less than 20–40 mesh presented little limitation on the hydrolysis rate of the biomass.46 Jacobsen and Wyman48 indicated that 5% (w/w) solid concentration gave the uniformity of the reaction system with negligible influences on the disruption of lignocellulosic biomass. Thus, 5% (w/w) solid concentration was employed in this study.
Fig. 1 showed that cellulose decomposed slowly at relatively low temperatures 140–160 °C. More than 80% and 55% cellulose residues were observed at 140 °C and 160 °C respectively even when the reaction time was prolonged to 60 min. It was found that the cellulose degradation was significantly enhanced as the temperature was elevated to 180 °C at which merely 12.1% cellulose residue remained after 60 min reaction. This abrupt increase in cellulose hydrolysis could be attributed to the disruption of crystallinity of cellulose. Further increasing reaction temperature to 200 °C and 220 °C resulted in the total dissolution of cellulose at 30 min and 10 min, respectively. Correspondingly, Xiang and colleagues29 reported that cellulose was susceptible to degradation when temperature was elevated to 215 °C or above in 0.07 (w/w)% H2SO4 solution. Sasaki et al.49 also found that cellulose was rapidly dissolved and depolymerized in supercritical water at temperatures ranging from 300–320 °C. Dissolution of cellulose during cellulose hydrolysis could result from a temperature-induced disruption of the hydrogen bond existing among the cellulose chains. In this regard, a parameter reflecting the effects of cellulose crystallinity as temperature varies appears essential to be incorporated into the kinetic model.
 |
| | Fig. 1 Yields of cellulose residue during the 1% (w/w) sulfuric acid hydrolysis of switchgrass at temperature of 140–220 °C. ◆: 140 °C; ■: 160 °C; ●: 180 °C; ▲: 200 °C; ★: 220 °C. | |
4.2. Release pattern of cellulose-derived glucose, 5-HMF and levulinic acid from switchgrass cellulose
4.2.1. Glucose production from switchgrass cellulose. Glucose yields from switchgrass cellulose were plotted with reaction time 0–60 min at 140–220 °C (Fig. 1). As shown in Fig. 2, at 140 °C, the glucose yield increased steadily over time, reaching the highest yield of 14.3% at 60 min. When temperature was increased from 140 °C to 160 °C, the glucose yield enhanced slowly over time, reaching the highest yield of 18.8% at 40 min, before declining gradually. The glucose yield at 180 °C presented a similar trend as that at 160 °C over the tested time course. However, results implied a higher rate constant of the formation and degradation of glucose at 180 °C. The maximum glucose yield of 31.5% was obtained at 15 min under 180 °C. The glucose yields obtained at elevated temperature (200 °C and 220 °C) presented much faster formation and degradation than those at lower temperatures (e.g. 180 °C). The peak yields of glucose obtained under 200 °C and 220 °C were 32.4% at 5 min and 42.8% at 1 min, respectively. These results revealed that relatively higher temperature (e.g. 220 °C) and limited reaction time (e.g. 1 min) was favored for the formation of glucose from cellulose compared to relatively lower temperatures (e.g. 140 °C) accompanied with prolonged time (e.g. 60 min). Lee et al.5 reported that the activation energy of cellulose hydrolysis is higher than that of glucose decomposition under dilute acid conditions at around 200 °C or below. Thus, a temperature increase could accelerate the rate constant of cellulose hydrolysis (k1) faster than glucose degradation rate constant (k2), thereby enhancing the glucose yield. Xiang et al.29 also demonstrated that elevated temperature (e.g. 215 °C) efficiently promoted cellulose hydrolysis and led to significant enhancement of glucose yield under acidic conditions (e.g. 0.07 (w/w)% H2SO4).
 |
| | Fig. 2 Yields of glucose during the 1% (w/w) sulfuric acid hydrolysis of switchgrass at temperature of 140–220 °C. ◆: 140 °C; ■: 160 °C; ●: 180 °C; ▲: 200 °C; ★: 220 °C. | |
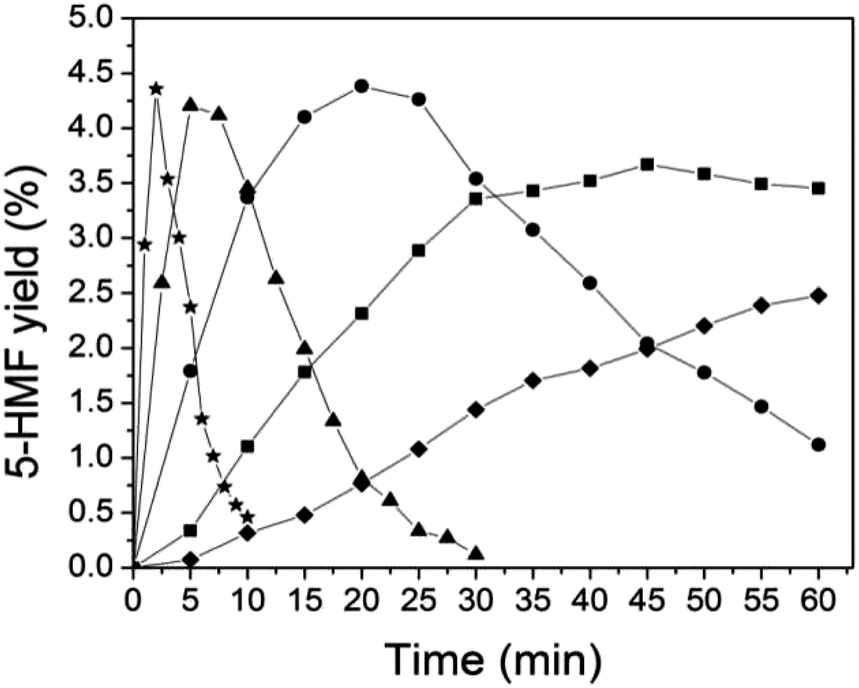
4.2.2. 5-HMF production from switchgrass derived-cellulose. The yields of 5-HMF as a function of reaction time at temperatures 140–220 °C are shown in Fig. 3. The formation of 5-HMF under tested conditions displayed similar trends as that of glucose, i.e. higher temperature with reduced reaction time led to higher 5-HMF yield. However, 5-HMF yields were very low under the tested conditions. The maximum 5-HMF yield obtained in this series of experiments was only 4.5% under 220 °C within 2 min. This indicated a high rate constant of 5-HMF degradation, which resulted in considerable 5-HMF degradation products (e.g. ∼60% levulinic acid) (Fig. 4). Many researchers reported that 5-HMF was not stable under acidic conditions. For example, Asghari and Yoshida50 reported that the dehydration of fructose by 0.03 M hydrochloric acid under 240 °C for 2 min resulted in low 5-HMF yields of 9.0% and 46.1% levulinic acid yield. Apart from the aforementioned degradation products from 5-HMF (e.g. levulinic acid), the low 5-HMF yield from lignocellulosic biomass (e.g. switchgrass) in acidic solution could also be due to the formation of ASL, which reacts with the glucose released from lignocellulosic biomass to form a complex called ASL–glucose,13 thus diminishing the possibility of the glucose conversion into 5-HMF. Although 5-HMF yield was low (less than 10%) with dilute acid (e.g. 1% (w/w) H2SO4), 5-HMF plays a key role as an important intermediate when understanding the mechanism of cellulose degradation under acidic conditions.8,9
 |
| | Fig. 3 Yields of 5-HMF during the 1% (w/w) sulfuric acid hydrolysis of switchgrass at temperature of 140–220 °C. ◆: 140 °C; ■: 160 °C; ●: 180 °C; ▲: 200 °C; ★: 220 °C. | |
 |
| | Fig. 4 Yields of levulinic acid during the 1% (w/w) sulfuric acid hydrolysis of switchgrass at temperature of 140–220 °C. ◆: 140 °C; ■: 160 °C; ●: 180 °C; ▲: 200 °C; ★: 220 °C. | |
4.2.3. Levulinic acid production from switchgrass derived-cellulose. Fig. 4 presents levulinic acid yields generated from switchgrass with 1% (w/w) H2SO4 at 140–220 °C over 0–60 min. It showed that at temperatures 140–180 °C, levulinic acid yield increased consistently over reaction time 0–60 min. The maximum levulinic acid yields were 4.2%, 23.8% and 59.5% for 140 °C, 160 °C and 180 °C at 60 min, respectively. When temperature was raised to 200 °C, levulinic acid yield increased significantly until the maximum yield of 60.4% was obtained at 30 min, then decreased slowly. Continuously increasing temperature to 220 °C resulted in a maximum levulinic acid yield of 57.8% within a relatively short reaction time of 10 min. Results indicated that the yield of levulinic acid under experimental conditions could hardly exceed 60%. Previous studies reported similar levulinic acid yield from cellulose with dilute acid. For example, Shen and Wyman8 reported that around 60% peaked levulinic acid yield was obtained with 0.927 M HCl under 180 °C or 200 °C, although levulinic acid showed a slow degradation rate constant. The limited levulinic acid production could be predominately attributed to the formation of humins observed by Patil and Lund.14 Humins formation could consume the 5-HMF yield thereby decrease the yield of levulinic acid to some extent.3,14 Results suggested that a considerable proportion of 5-HMF condensed to humins and led to the limited yield of levulinic acid observed under dilute acid conditions.
4.3. Kinetic model of cellulose degradation
4.3.1. Determination and comparison of the kinetic rate constants from both Model I and Model II. The conventional model (Model I) and modified model (Model II) described in Section 2 were applied to fit the experimental data obtained for glucose, 5-HMF and levulinic acid generated from switchgrass derived-cellulose. Fig. 5a–d presents the fitted curves for yields of cellulose, glucose, 5-HMF and levulinic acid from both conventional model (Model I) and modified model (Model II), respectively. Results showed that the modified model (Model II) fitted the experimental data marginally better than the conventional model (Model I). This is expected because Model II introduced more parameters into the system, and hence extra degrees of freedom into the parameter fitting. The coefficient of determination, R2, for the yields of cellulose residue, glucose, 5-HMF and levulinic acid based on Model I and Model II (Tables 1 and 2) ranged from 0.92–0.99 and 0.97–0.99, respectively, which suggested that both two models fitted the experimental data well.
 |
| | Fig. 5 Fitted curves of cellulose residue yield (a), glucose yield (b), 5-HMF yield (c) and levulinic acid yield (d) vs. time based on conventional kinetic model (Model I) and modified model (Model II) during 1% (w/w) sulfuric acid hydrolysis of switchgrass at temperatures of 140–220 °C. ○: 140 °C; +: 160 °C; △: 180 °C; *: 200 °C; □: 220 °C. Red dash line: Model I; blue line: Model II. | |
Table 1 Kinetic constants for the conventional kinetic model (Model I)
| Temperature (°C) |
Conventional model (Model I) |
| |
R2 |
| k1 (min−1) |
k2 (min−1) |
k3 (min−1) |
k′3 (min−1) |
k4 (min−1) |
Cellulose |
Glucose |
5-HMF |
Levulinic acid |
| 140 |
0.0043 |
0.0136 |
0.0652 |
0.0578 |
0.0009 |
0.963 |
0.985 |
0.989 |
0.985 |
| 160 |
0.0106 |
0.0336 |
0.1649 |
0.1596 |
0.0013 |
0.979 |
0.980 |
0.982 |
0.995 |
| 180 |
0.0459 |
0.0706 |
0.4991 |
0.4617 |
0.0092 |
0.987 |
0.978 |
0.977 |
0.995 |
| 200 |
0.1836 |
0.2036 |
1.6883 |
1.3503 |
0.0133 |
0.998 |
0.976 |
0.986 |
0.965 |
| 220 |
0.6779 |
0.4510 |
4.7858 |
3.4327 |
0.0181 |
0.994 |
0.973 |
0.925 |
0.918 |
Table 2 Kinetic constants and parameters α, β, γ for the modified kinetic model (Model II)
| Temperature (°C) |
Modified model (Model II) |
| |
R2 |
| k1 (min−1) |
k2 (min−1) |
k3 (min−1) |
k4 (min−1) |
α |
β |
γ |
Cellulose |
Glucose |
5-HMF |
Levulinic acid |
| 140 |
0.0210 |
0.0111 |
0.0570 |
0 |
0.7294 |
0 |
0 |
0.992 |
0.983 |
0.996 |
0.981 |
| 160 |
0.0249 |
0.0299 |
0.1406 |
0.0006 |
0.4423 |
0.0639 |
0 |
0.996 |
0.986 |
0.998 |
0.993 |
| 180 |
0.0602 |
0.0643 |
0.3927 |
0.0019 |
0.1177 |
0.1682 |
0.079 |
0.995 |
0.996 |
0.996 |
0.997 |
| 200 |
0.1853 |
0.2031 |
1.3754 |
0.0086 |
0.0030 |
0.1835 |
0.1176 |
0.998 |
0.978 |
0.998 |
0.998 |
| 220 |
0.6810 |
0.4502 |
3.7774 |
0.0110 |
0.0016 |
0.2089 |
0.2310 |
0.994 |
0.979 |
0.990 |
0.986 |
Tables 1 and 2 also present the values of the estimated parameters for the conventional model (Model I) and the modified model (Model II), respectively. At relatively low temperatures (140–180 °C), the values of rate constant k1 determined from the conventional model (Model I) were lower than those determined from the modified model (Model II). Nevertheless, when temperature was raised to 200 °C and 220 °C, the values of k1 obtained from both Model I and Model II were analogous. For example, the value of k1 for Model I was 0.0043 min−1 at 140 °C, compared to 0.0210 min−1 for Model II at identical temperature, while both values of k1 obtained from Model I and Model II increased to 0.1836 min−1 and 0.1853 min−1 at 200 °C, respectively. The relatively higher value of rate constant of cellulose hydrolysis obtained based on Model II was the consequence of the incorporation of parameter α, representing the physical barriers of cellulose (crystallinity of cellulose). In this regard, the cellulose degradation rate constant (k1) obtained based on Model II could particularly reflect the reaction chemistry without the interference of physical limitations (e.g. crystallinity). It was noticeable that the value of k1 for Model I was similar to that for Model II as temperature was elevated to 200 °C or above, indicating the reduction of the crystalline region of cellulose at such temperature levels.
Rate constants k2, k3 and k4 obtained from Model II were lower than those from Model I under the tested conditions. This is the consequence of the introduction of the additional parameters (α, β and γ) representing the crystallinity of cellulose and undesired byproducts (e.g. ASL–glucose and humins) in Model II, which implied that only partial cellulose, glucose and 5-HMF were decomposed to downstream desired byproducts, thereby obtaining the relatively lower degradation rate constants.
Cellulose, as well as its derivatives including glucose, 5-HMF and levulinic acid, displayed significantly different degradation rate constants. The rate constant of glucose degradation (k2) was higher than that of cellulose hydrolysis (k1) at low temperature (e.g. 140–160 °C) for Model I, where the ratio of k2/k1 was approximately 3.0. In comparison, the ratio of k2/k1 for Model II was 0.5–1.2 over an identical temperature range (e.g. 140–160 °C), indicating that without the physical obstacle caused by the hydrogen bonding in cellulose, the hydrolysis rate constant of cellulose could be similar to that of glucose. Elevating the temperature to 200 °C or above resulted in the significant increase in k1. Although k2 was enhanced as well, it increased more slowly than k1, thereby leading to the decrease of the ratio of k2/k1 to around 0.6–1.0 for both Model I and Model II, which implied that higher temperature was favorable for glucose production.
The degradation rate constant of 5-HMF (k3) was especially higher than that of its precursor glucose (k2) obtained from either Model I or Model II. The ratio of k3/k2 is 4.8–10.6 for Model I, and 4.7–8.4 for Model II over the tested temperatures (i.e. 140–220 °C). This result further corroborated the observations reported in previous studies that 5-HMF is susceptible to degradation under acid conditions.51,52 It was also observed that the decomposition rate constant of levulinic acid (k4) was significantly lower than that of its formation (i.e. k′3 for Model I and k3 for Model II). The ratio of k4/k′3 for Model I was merely 0.005–0.020 and the ratio of k4/k3 for Model II was 0.004–0.006 under the temperature range of 140–220 °C, respectively. However, the yield of levulinic acid obtained in this study didn't exceed 60% (Section 4.2.3) although it showed stability under acidic conditions.8 In this regard, the consumption of levulinic acid precursors i.e. glucose and 5-HMF to undesired products like ASL–glucose and humins could play a predominant role in determining the production of levulinic acid.
4.3.2. Determination of parameters α, β and γ from Model II. The modified model (Model II) appeared well fit in describing the acid-catalyzed cellulose degradation than the conventional model (Model I) due to the addition of the parameters α, β and γ (Section 4.2). Fig. 6 shows the relationship between these parameters (α, β and γ) and the temperatures employed in this set of experiments (140–220 °C). Each of the three parameters exhibited different temperature dependency. It was found that the value of α was 72.9% at 140 °C, implying that a considerable crystalline region of cellulose exists at this temperature level, thereby impeding the hydrolysis of cellulose to some extent. The value of α decreased as reaction temperature increased, declining to 44.2% and 11.8% at 160 °C and 180 °C, respectively. As the temperature further raised to 200 °C or above, the value of α was significantly reduced to a negligible value, which indicated that the majority of crystalline cellulose was transformed to an amorphous structure, thus acid could easily penetrate into the cellulose and catalyze the cleavage of the β-1,4-glycosidic bonds. This result corroborated the findings in previous studies, i.e. there is a “breaking temperature” for the disruption of crystalline cellulose.29,49,53,54 For example, Xiang et al. used 0.07% (w/w) H2SO4 to hydrolyze cellulose under 185–245 °C, and found that the rate constant of hydrolysis reaction of cellulose suddenly increased when temperature reached 215 °C, indicating that the elimination of the hydrogen bonding of cellulose took place around 215 °C.29 Agarwal et al.53 applied infrared (IR) spectra observing the hydrogen bond changing in cellulose over temperatures ranging from 27–277 °C. It was observed that cellulose was dominated with inter chain hydrogen bonds at low temperatures (27–127 °C). When the temperature was elevated to 177–277 °C, considerable hydrogen bonds transformed to longer, weaker inter-chain ones.
 |
| | Fig. 6 Values of α, β and γ vs. temperatures at 140 °C, 160 °C, 180 °C, 200 °C and 220° C. | |
Apart from parameter α, the value of β and γ were plotted with temperatures ranging from 140–220 °C as well. The value of β was negligible at 140 °C, suggesting few undesired products (e.g. ASL–glucose) formed at low temperature. Apart from few ASL and glucose generated at lower temperatures, the possible explanation could be that the relatively low temperature resulted in less hydronium ions55 causing few protonized glucose to condense with nucleophilic reaction partners (e.g. ASL) (Section 2.2),11,13,31 thereby leading to negligible formation of undesired products (e.g. ASL–glucose). As temperature was elevated, the value of β increased progressively and reached the maximum value 20.9% at 220 °C. This result indicated that the value of β is temperature dependent. Relatively high temperatures could result in higher production of glucose derived undesired products (e.g. ASL–glucose), thereby reducing the yield of 5-HMF generated from the subsequent glucose degradation. Previous researches also reported that the interaction between the glucose and ASL was prone to be severe at relatively high temperatures. For example, Xiang et al.11 compared the glucose degradation under water-only conditions with and without lignin at 200 °C using 0.1% (w/w) sulfuric acid over 5 min. The glucose degradation rate constant in the medium with lignin was around 2 times faster than that without lignin.
The values of γ at low temperatures (i.e. 140–160 °C) were imperceptible, which implied few undesired products (e.g. humins) were generated over this range of temperatures during the tested reaction time (0–60 min). Patil and Lund14 also observed indistinct formation of humins with the assistance of scanning electron microscopy (SEM) when treating 5-HMF with 1% (w/w) H2SO4 at relatively low temperature (135 °C) for 22 min. However, the formation of humins was detected as reaction time was prolonged to 120 min, suggesting that reaction time could affect the production of humins to some extent. It was found that the value of γ increased abruptly to 7.9% when temperature reached 180 °C. As temperature further increased to 200 °C and 220 °C, the value of γ reached 11.8% and 23.1%, respectively. This finding was coincident with the statements proposed by previous studies, i.e. 5-HMF is prone to form other products (e.g. humins) rather than levulinic acid under acidic conditions when the temperature is increased,56,57 thus potentially limiting the production of levulinic acid. Levulinic acid was a stable compound with relatively low degradation rate constant even at high temperature (e.g. 0.0086 min−1 at 200 °C) in 1% (w/w) H2SO4 solution (Table 2). Thus, the formation of parallel product (e.g. humins) is distinguished as the key negative factor in determining the yield of levulinic acid.
Results showed that the value of α was inversely proportional to temperature, thereby higher temperatures with limited α value were favored for the elimination of the crystallinity of cellulose, and the subsequent production of glucose, 5-HMF and levulinic acid. In contrast, the values of β and γ were directly proportional to temperature, which suggested that lower temperatures could be recommended for the reduced production of undesired products (e.g. ASL–glucose and humins), thus favored for the generation of glucose, 5-HMF and levulinic acid.
4.4. Sensitivity analysis of parameters obtained in Model II
The parameters, i.e. α, β and γ, incorporated in Model II representing the effects of crystalline cellulose, as well as undesired products (e.g. ASL–glucose and humins), all had negative impacts on the production of cellulose-derived glucose, 5-HMF and levulinic acid. A sensitivity analysis of the impact of rate constants (k1, k2, k3 and k4) as well as these parameters (α, β and γ) upon the production of glucose, 5-HMF and levulinic acid predicted in Model II is necessary in order to enhance the understanding of the production of these value-added compounds. Table 3 presents a comparison of the sensitivity of the peaked yields of glucose, 5-HMF and levulinic acid generated by Model II to small changes in the reaction rate constants and the parameters α, β and γ under tested temperatures (140–220 °C). In this table, variations in the maximum yields of glucose, 5-HMF and levulinic acid were recorded as a result of changing these parameters by ±10%.
Table 3 The sensitivity of the peaked yield of glucose, 5-HMF and levulinic acid to the parameters α, β, γ and rate constantsa
| Change in parameters |
Change in glucose yield (%) |
Change in 5-HMF yield (%) |
Change in levulinic acid yield (%) |
| T1 |
T2 |
T3 |
T4 |
T5 |
T1 |
T2 |
T3 |
T4 |
T5 |
T1 |
T2 |
T3 |
T4 |
T5 |
| T1: 140 °C; T2: 160 °C; T3: 180 °C; T4: 200 °C; T5: 220 °C. |
| +10% k1 |
+0.53 |
+1.01 |
+1.57 |
+1.83 |
+1.49 |
+0.12 |
+0.17 |
+0.17 |
+0.20 |
+0.23 |
+0.28 |
+1.29 |
+1.30 |
+0.53 |
+0.27 |
| −10% k1 |
−0.62 |
−1.09 |
−1.53 |
−1.78 |
−1.61 |
−0.13 |
−0.18 |
−0.19 |
−0.22 |
−0.23 |
−0.28 |
−1.32 |
−1.30 |
−0.58 |
−0.25 |
| +10% k2 |
−0.47 |
−0.92 |
−1.51 |
−1.69 |
−2.08 |
+0.15 |
+0.17 |
+0.19 |
+0.22 |
+0.23 |
+0.31 |
+1.09 |
+0.96 |
+0.42 |
+0.40 |
| −10% k2 |
+0.49 |
+0.94 |
+1.63 |
+1.72 |
+2.21 |
−0.16 |
−0.18 |
−0.20 |
−0.23 |
−0.24 |
−0.38 |
−1.13 |
−1.01 |
−0.43 |
−0.44 |
| +10% k3 |
— |
— |
— |
— |
— |
−0.16 |
−0.35 |
−0.39 |
−0.43 |
−0.45 |
+0.16 |
+0.31 |
+0.08 |
+0.01 |
+0.01 |
| −10% k3 |
— |
— |
— |
— |
— |
+0.18 |
+0.37 |
+0.41 |
+0.43 |
+0.47 |
−0.18 |
−0.35 |
−0.09 |
−0.01 |
−0.01 |
| +10% k4 |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
0 |
−0.02 |
−0.38 |
−0.79 |
−0.80 |
| −10% k4 |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
0 |
+0.03 |
+0.39 |
+0.80 |
+0.78 |
| +10% α |
−3.55 |
−1.49 |
−0.43 |
0 |
0 |
−0.59 |
−0.30 |
−0.08 |
0 |
0 |
−1.34 |
−1.22 |
−0.81 |
0 |
0 |
| −10% α |
+3.55 |
+1.48 |
+0.43 |
0 |
0 |
+0.61 |
+0.28 |
+0.08 |
0 |
0 |
+1.37 |
+1.18 |
+0.79 |
0 |
0 |
| +10% β |
— |
— |
— |
— |
— |
0 |
−0.02 |
−0.08 |
−0.15 |
−0.45 |
0 |
−0.14 |
−1.21 |
−1.72 |
−2.08 |
| −10% β |
— |
— |
— |
— |
— |
0 |
+0.02 |
+0.08 |
+0.13 |
+0.47 |
0 |
+0.15 |
+1.21 |
+1.78 |
+2.03 |
| +10% γ |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
0 |
0 |
−0.05 |
−1.28 |
−2.68 |
| −10% γ |
— |
— |
— |
— |
— |
— |
— |
— |
— |
— |
0 |
0 |
+0.05 |
+1.30 |
+2.73 |
The glucose yield was related to k1, k2 and α. It was found that the ±10% change of the values of k1 and k2 over the tested temperatures (140–220 °C) resulted in the alteration of maximum glucose yield by around ±0.5% to ±1.5% and ±0.5% to ±2.0%, respectively. By contrast, altering the α parameter by ±10% modified the resulting maximum glucose yield by around ±0.4% to ±3.5% throughout 140–180 °C. This result indicated that the glucose yield was more sensitive to variations in the parameter α than the rate constants k1 and k2, and hence α could be considered as the key factor in controlling the production of glucose under these experimental conditions. However, at elevated temperatures (200 °C and 220 °C), because the value of α was almost negligible, its influence on glucose yield declined to an imperceptible level, thus the glucose production could be predominately controlled by the rate constants k1 and k2. The rate constants k1, k2 and k3 as well as α and β codetermined the 5-HMF yield. Variations in these parameters led to small changes in 5-HMF yield (around 0–±0.6%), which suggested that no factor was compelling in determining the yields of 5-HMF under tested conditions, thereby demonstrating the potential complexity of improving the production of 5-HMF. Changes in all of the parameters in Model II (k1, k2, k3, k4, α, β and γ) impacted upon the levulinic acid yields. A ±10% change in values of rate constant (k1, k2, k3 and k4) resulted in merely 0–±1.3% alteration of levulinic acid yield. Comparatively, ±10% alteration of α, β and γ led to 0–±1.4%, 0–±2.1% and ±0.01–±2.7% variation in levulinic acid yields respectively under the tested temperatures. Results demonstrated that levulinic acid production was more sensitive to variations in the parameters α than other additional parameters (β and γ) and the rate constants (k1, k2, k3 and k4) under lower temperatures (e.g. 140 °C). The sensitivity of the levulinic acid yield to small changes in the additional parameters (e.g. α) became less significant compared with rate constants (e.g. k1 and k2) when temperature was elevated to 160–180 °C. However, when the temperature reached 200–220 °C, the β and γ parameters had the most pronounced impact upon the yield of levulinic acid among all the parameters used in Model II. For example, at 220 °C, changing γ by ±10% resulted in the ±2.7% alteration of levulinic acid yield. In this regard, due to the significant impact of parameters β and γ at higher temperatures (200–220 °C) as well as the apparent influence of parameter α under lower temperatures (e.g. 140 °C) on the production of levulinic acid, medium temperatures, such as 160–180 °C, can be recommended to minimize the negative influence of these parameters (i.e. α, β and γ) on the levulinic acid generation, thereby optimizing its yield. This deduction was demonstrated by the results based on Model II. For example, the maximum levulinic acid yield predicted based on Model II was 63.6% at 160 °C over 90 min, which was higher than that obtained under 200 °C (57.7%).
5. Conclusion
The production of glucose, 5-HMF and levulinic acid from switchgrass cellulose was investigated at temperatures of 140–220 °C for 0–60 min with 1% (w/w) H2SO4 in batch tubular reactors. The highest glucose yield of 42.8% was observed under experimental conditions. Results suggested that elevated temperature and reduced reaction time was favorable for the formation of glucose. The levulinic acid yield did not exceed 60% under experimental conditions although its degradation rate constant was relatively low at tested temperatures. 5-HMF appeared unstable under dilute acid conditions with the highest yield of less than 5%. A comprehensive mechanistic kinetic model was developed to interpret the release pattern of glucose, 5-HMF and levulinic acid from switchgrass cellulose. This novel model incorporating the parameters α reflecting the effect of cellulose crystallinity as well as the parameters β and γ representing the influence of undesired products from glucose and 5-HMF, including ASL–glucose and humins, inferred some quantitative estimation of the reduced yields of glucose, 5-HMF and levulinic acid due to these undesired reaction limitations. Results showed that this model fitted experimental data of yields of glucose, 5-HMF and levulinic acid from cellulose with R2 ≥ 0.97. Sensitivity analysis of these parameters indicated that the glucose yield was more sensitive to changes in the value of the α parameter than the rate constants at low temperatures (140–180 °C). The value of α became negligible as the temperature was raised to 200–220 °C, therefore causing rapid increase in cellulose hydrolysis and glucose yield. The levulinic acid yield was also sensitive to the alterations in the value of α parameter under low temperatures (e.g. 140 °C) whereas β and γ parameters were the predominant controlling factors in determining the yield of levulinic acid at high temperatures (e.g. 200–220 °C). Effects of the three parameters (α, β and γ) on levulinic acid yields were reduced at medium temperatures (160–180 °C). It suggested that such temperature benefitted levulinic acid production. None of the three parameters or the rate constants was found to be particularly compelling in effecting the 5-HMF yields. Our results suggested that this new model possessed the capability in predicting the generation of glucose, 5-HMF and levulinic acid from lignocellulosic biomass derived cellulose under dilute acid batch pretreatment conditions. Although accurate measurement of cellulose crystallinity, ASL–glucose and humins has been proven very difficult, this kinetic model provided new insights into the possible mechanisms of cellulose degradation that cellulose crystallinity and formation of ASL–glucose and humins could be among the key factors that govern the yields of products of interest. However, crystallinity index of cellulose, ASL–glucose and humins predicted by this model were not able to be compared to experimental measurements because few technologies exist for these undesired reaction limitations to be accurately quantified. In addition, this model could prove less accurate at predicting the hydrolysis of cellulose with high substrate concentration because it didn't incorporate parameters reflecting the heterogeneous properties of cellulose.
Acknowledgements
We are grateful for the support from the Department of Biological Systems Engineering and the Bioproducts, Sciences and Engineering Laboratory at Washington State University. The material in this work was also supported by DARPA YFA Award # N66001-11-1-4141/P00001. The authors also thank Drs Qian Xiang and Ian O'Hara for their kind review of this paper and helpful suggestions.
References
- G. W. Huber, J. N. Chheda, C. J. Barrett and J. A. Dumesic, Science, 2005, 308, 1446–1450 CrossRef CAS PubMed.
- B. Yang and C. E. Wyman, Biofuels, Bioprod. Biorefin., 2008, 2, 26–40 CrossRef CAS.
- D. W. Rackemann and W. O. S. Doherty, Biofuels, Bioprod. Biorefin., 2011, 5, 198–214 CrossRef CAS.
- A. S. Mamman, J.-M. Lee, Y.-C. Kim, I. T. Hwang, N.-J. Park, Y. K. Hwang, J.-S. Chang and J.-S. Hwang, Biofuels, Bioprod. Biorefin., 2008, 2, 438–454 CrossRef CAS.
- Y. Y. Lee, P. Iyer and R. W. Torget, Adv. Biochem. Eng./Biotechnol., 1999, 65, 94–115 CrossRef.
- R. W. Torget, J. S. Kim and Y. Y. Lee, Ind. Eng. Chem. Res., 2000, 39, 2817–2825 CrossRef CAS.
- R. W. H. Torget, T. K. Hayward and R. Elander, 19th Symposium on Biotechnology for Fuels and Chemicals, Colorado Springs, 1997 Search PubMed.
- J. Shen and C. E. Wyman, AIChE J., 2012, 58, 236–246 CrossRef CAS.
- B. Girisuta, L. P. B. M. Janssen and H. J. Heeres, Ind. Eng. Chem. Res., 2007, 46, 1696–1708 CrossRef CAS.
- J. Horvat, B. Klaić, B. Metelko and J. Vitomir, Tetrahedron Lett., 1985, 26, 2111–2114 CrossRef CAS.
- Q. Xiang, J. S. Kim and Y. Y. Lee, Appl. Biochem. Biotechnol., 2003, 105–108, 337–352 CrossRef CAS.
- H. L. Trajano and C. E. Wyman, in Aqueous Pretreatment of Plant Biomass for Biological and Chemical Conversion to Fuels and Chemicals, ed. C. E. Wyman, John Wiley&Sons, Ltd., West Sussex, 2013, ch. 6, pp. 103–123 Search PubMed.
- Q. Xiang, Y. Y. Lee and R. W. Torget, Appl. Biochem. Biotechnol., 2004, 113–116, 1127–1138 CrossRef CAS.
- S. K. R. Patil and C. R. F. Lund, Energy Fuels, 2011, 25, 4745–4755 CrossRef CAS.
- O. A. Battista, Ind. Eng. Chem. Res., 1950, 42, 502–507 CrossRef CAS.
- S. Park, J. O. Baker, M. E. Himmel, P. A. Parilla and D. K. Johnson, Biotechnol. Biofuels, 2010, 3, 10–19 CrossRef PubMed.
- N. Mosier, in Aqueous Pretreatment of Plant Biomass for Biological and Chemical Conversion to Fuels and Chemicals, ed. C. E. Wyman, John Wiley&Sons, Ltd., West Sussex, 2013, ch. 7, pp. 129–140 Search PubMed.
- B. Yang, M. C. Gray, C. Liu, T. A. Lloyd, S. L. Stuhler, A. O. Converse and C. E. Wyman, ACS Symp. Ser., 2004, 889, 100–125 CrossRef CAS PubMed.
- J. F. Saeman, Ind. Eng. Chem. Res., 1945, 37, 43–52 CrossRef CAS.
- S. W. McKibbins, J. F. Harris, J. F. Saeman and W. K. Neill, For. Prod. J., 1962, 12, 17–23 CAS.
- R. D. Fagan, H. E. Grethlein, A. O. Converse and A. Porteous, Environ. Sci. Technol., 1971, 5, 545–547 CrossRef CAS.
- N. S. Mosier, C. M. Ladisch and M. R. Ladisch, Biotechnol. Bioeng., 2002, 79, 610–618 CrossRef CAS PubMed.
- R. Aguilar, J. A. Ramirez, G. Garrote and M. Vazquez, J. Food Eng., 2002, 55, 309–318 CrossRef.
- A. M. Orozco, A. H. Al-Muhtaseb, A. B. Albadarin, D. Rooney, G. M. Walker and M. N. M. Ahmad, RSC Adv., 2011, 1, 839–846 RSC.
- Q. Yu, X. Zhuang, Q. Wang, W. Qi, X. Tan and Z. Yuan, Bioresour. Technol., 2012, 116, 220–225 CrossRef CAS PubMed.
- Y. Lu and N. S. Mosier, Biotechnol. Bioeng., 2008, 101, 1170–1181 CrossRef CAS PubMed.
- G. Garrote, H. Domínguez and J. C. Parajo, Process Biochem., 2001, 36, 571–578 CrossRef CAS.
- A. Mittal, S. G. Chatterjee, G. M. Scott and T. E. Amidon, Chem. Eng. Sci., 2009, 64, 3031–3041 CrossRef CAS PubMed.
- Q. Xiang, Y. Y. Lee, P. O. Pettersson and R. W. Torget, Appl. Biochem. Biotechnol., 2003, 107, 505–514 CrossRef.
- G. Brodeur, E. Yau, K. Badal, J. Collier, K. B. Ramachandran and S. Ramakrishnan, Enzyme Res., 2011, 2011, 787532–787548 Search PubMed.
- D. Fengel and G. Wegener, in Wood—Chemistry, Ultrastructure, Reactions, ed. D. Fengel and G. Wegener, Walter de Gruyter, Berlin and New York, 1984, pp. 279–281 Search PubMed.
- G. C. A. Luijkx, V. F. Rantwijk and V. B. Herman, Carbohydr. Res., 1993, 242, 131–139 CrossRef CAS.
- B. Girisuta, L. P. B. M. Janssen and H. J. Heeres, Green Chem., 2006, 8, 701–709 RSC.
- C. C. Conrad and A. G. Scroggie, Ind. Eng. Chem. Res., 1945, 37, 592–598 CrossRef CAS.
- Y. Ma, X. Zhao, H. Zhang and Z. Wang, Ind. Crops Prod., 2011, 33, 403–408 CrossRef CAS PubMed.
- C. H. Stephens, P. M. Whitmore, H. R. Morris and M. E. Bier, Biomacromolecules, 2008, 9, 1093–1099 CrossRef CAS PubMed.
- P. Petchpradab, T. Yoshida, T. Charinpanitkul and Y. Matsumura, Ind. Eng. Chem. Res., 2009, 48, 4587–4591 CrossRef CAS.
- W. S. Mok and M. J. Antal, Ind. Eng. Chem. Res., 1992, 31, 94–100 CrossRef CAS.
- B. C. Saha, J. Ind. Microbiol. Biotechnol., 2003, 30, 279–291 CrossRef CAS PubMed.
- P. R. Adler, M. A. Sanderson, A. A. Boateng, P. J. Weimer and H.-J. G. Jung, Agron J., 2006, 98, 1518–1525 CrossRef CAS.
- K. David and A. J. Ragauskas, Energy Environ. Sci., 2010, 3, 1182–1190 CAS.
- C. E. Wyman, B. E. Dale, V. Balan, R. T. Elander, M. T. Holtzapple, R. S. Ramirez, M. R. Ladisch, N. S. Mosier, Y. Y. Lee, R. Gupta, S. R. Thomas, B. R. Hames, R. Warner and R. Kumar, in Aqueous Pretreatment of Plant Biomass for Biological and Chemical Conversion to Fuels and Chemicals, ed. C. E. Wyman, John Wiley&Sons, Ltd., West Sussex, 2013, ch. 12, pp. 239–258 Search PubMed.
- K. Lindsey, A. Johnson, P. Kim, S. Jackson and N. Labbé, Biomass Bioenergy, 2013, 56, 29–37 CrossRef CAS PubMed.
- A. Sluiter, B. Hames, R. Ruiz, C. Scarlata, J. Sluiter, D. Templeton and D. Crocker, Technical Report. NREL/TP-510-42618, 2011.
- L. Yan, D. D. Laskar, S.-J. Lee and B. Yang, RSC Adv., 2013, 3, 24090–24098 RSC.
- V. S. Chang and M. T. Holtzapple, Appl. Biochem. Biotechnol., 2000, 84–86, 5–37 CrossRef CAS.
- C. E. Wyman, B. E. Dale, R. T. Elander, M. Holtzapple, M. R. Ladisch and Y. Y. Lee, Bioresour. Technol., 2005, 96, 1959–1966 CrossRef CAS PubMed.
- S. E. Jacobsen and C. E. Wyman, Ind. Eng. Chem. Res., 2002, 41, 1454–1461 CrossRef CAS.
- M. Sasaki, Z. Fang, Y. Fukushima, T. Adschiri and K. Arai, Ind. Eng. Chem. Res., 2000, 39, 2883–2890 CrossRef CAS.
- F. S. Asghari and H. Yoshida, Ind. Eng. Chem. Res., 2006, 45, 2163–2173 CrossRef CAS.
- G. W. Huber and J. A. Dumesic, Catal. Today, 2006, 111, 119–132 CrossRef CAS PubMed.
- Y. Roman-Leshkov, C. J. Barrett, Z. Y. Liu and J. A. Dumesic, Nature, 2007, 447, 982–985 CrossRef CAS PubMed.
- V. Agarwal, G. W. Huber, W. C. Conner, Jr and S. M. Auerbach, J. Chem. Phys., 2011, 135, 134506–134518 CrossRef PubMed.
- R. Hashaikeh, Z. Fang, I. S. Butler, J. Hawari and J. A. Kozinski, Fuel, 2007, 86, 1614–1622 CrossRef CAS PubMed.
- S. Kumar, R. Gupta, Y. Y. Lee and R. B. Gupta, Bioresour. Technol., 2010, 101, 1337–1347 CrossRef CAS PubMed.
- M.-M. Titirici, M. Antonietti and N. Baccile, Green Chem., 2008, 10, 1204–1212 RSC.
- C. Yao, Y. Shin, L. Wang, C. F. Windisch, W. D. Samuels, B. W. Arey, C. Wang, W. M. Risen and G. J. Exarhos, J. Phys. Chem., 2007, 111, 15141–15145 CAS.
|
| This journal is © The Royal Society of Chemistry 2014 |
Click here to see how this site uses Cookies. View our privacy policy here.