Toxicity of various kinds of ionic liquids towards the cell growth and end product formation of the probiotic strain, Propionibacterium freudenreichii†
Hamidreza Hajfarajollah,
Babak Mokhtarani*,
Ali sharifi,
Mojtaba Mirzaei and
Ali Afaghi
Chemistry and Chemical Engineering Research Center of Iran, P.O. Box 14335-186, Tehran, Iran. E-mail: mokhtaranib@ccerci.ac.ir; Fax: +98 2144580781; Tel: +98 2144580770
First published on 26th February 2014
Abstract
This paper reports on an investigation into the toxicity of various kinds of 1-alkyl-3-methylimidazolium ionic liquids (ILs) including [BMIM][BF4], [HMIM][BF4], [BMIM][PTS], [HMIM][PTS], [BMIM][NO3], [HMIM][NO3], [OMIM][NO3], [BMIM][MeOSO3], [OMIM][MeOSO3], [BMIM][SCN], [OMIM][SCN], towards the cell growth of a probiotic strain Propionibacterium freudenreichii subsp. freudenreichii; in addition, the effects of ILs on the production of intracellular vitamin B12 (VB12) and extracellular propionic acid (PA) by the mentioned strain were investigated for the first time. The results of the study demonstrated that all ILs were toxic towards the microorganism in concentrations above 1% (w/v). The most toxic IL among them, however, was [OMIM][NO3] which inhibited the cell growth at even low concentrations of 0.005% (w/v). The inhibitory concentration after 48 h (IC50-48![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) h) for this IL was 0.005% (w/v). The least toxic IL was [BMIM][PTS] with IC50-48h corresponding to 1% (w/v). Production of PA was related to the concentration of IL. However, a complex relation between production of intracellular product, VB12, and the amount of IL in the broth was observed. The information in this paper could be useful for researchers who work in the fields of extractive fermentation, ionic liquid extraction and environmental pollution.
h) for this IL was 0.005% (w/v). The least toxic IL was [BMIM][PTS] with IC50-48h corresponding to 1% (w/v). Production of PA was related to the concentration of IL. However, a complex relation between production of intracellular product, VB12, and the amount of IL in the broth was observed. The information in this paper could be useful for researchers who work in the fields of extractive fermentation, ionic liquid extraction and environmental pollution.
Introduction
Ionic Liquids (ILs) are organic salts which remain as liquids under ambient temperatures.1 They normally consist of an organic cation (e.g. imidazolium, pyridinium), and a polyatomic inorganic anion (e.g. tetrafluoroborate, hexafluorophosphate).2 ILs have a number of interesting properties, such as high polarity, ionic conductivity, and excellent chemical stability. They are also nonvolatile, nonflammable, and thermally very stable.3 These properties make them excellent solvents for a wide range of inorganic and organic materials. Therefore, they have been employed successfully in catalytic synthesis,4 analytical chemistry,5 coordination chemistry,6 polymer materials,7 fluorine chemistry8 and nanotechnology.9 Since ILs introduced for replacing traditional organic solvents, investigation on the toxicity of them is of prominent importance. Recent studies have shown that the toxicity of many ionic liquids are similar to those of the industrial solvents that they may replace.10,11Application of ILs in the field of fermentation and biotechnology is being expanded. Applications in these areas can be divided into two main branches: enzymatic and whole-cell processes. The former applications are the most abundant in the researches. Increased activity and stability of several enzymes in the presence of ILs have been reported. Some examples of successful enzymatic catalysis in the presence of ILs are perhydrolysis, transesterification and ammoniolysis reactions.12 The latter applications are still scarce in the literature and are restricted to microorganisms able to tolerate the presence of ILs.10,11 The use of ILs in whole-cell processes has been mainly focused to the so called “extractive fermentation”. The extractive fermentation technology is based on the addition of a second liquid phase (immiscible in water) for the in situ removal of toxic metabolic products in order to avoid cell inhibition.13 Other examples in which the IL may be in a direct contact with the microorganisms are liquid–liquid extraction of biomaterials using ILs14 and lignocellulosic substrates pretreatment.15
Lignocellulosic materials, such as wood wastes, waste paper and agricultural residues have been recognized as a potential sustainable source of sugars.16 To enhance the enzymatic or microbial digestibility of these polysaccharide components, pretreatment of lignocellulose with ILs could be performed.16 However, since after pretreatment, ILs may remain and introduce to the downstream processes such as fermentation, they can negatively impact these processes.17 Therefore, the toxicity of ILs on bacteria should be extensively investigated.
Probiotic bacteria like Propionibacterium freudenreichii are considered GRAS (generally recognized as safe) with applications in food and pharmaceutical industries.18 P. freudenreichii produces vitamin B12 (VB12) intracellularly and propionic acid (PA) and acetic acid (AA) extracellularly. Propionic acid has an inhibitory effect on the cell growth and end products formation.19 Therefore, there are methods to simultaneous removal of acids from fermentation broth to enhance VB12 production. One of them is extractive fermentation by which the acid can be simultaneously removed by a solvent from fermentation broth. Gu et al.20 used three kinds of traditional solvents for this purpose. The solvents were trilaurylamine in 2-octanol, 1-dodecanol, and oleyl alcohol. Among them, the solvent containing 2-octanol showed the best acid extraction performance, however it was also toxic to Propionibacteria. ILs are emerging as alternative solvents for organic compounds traditionally used in extraction. Hence, extractive fermentation for acid removal with ILs is being investigated.19 The toxicity of ILs is however, a major drawback for further application of ILs in extractive fermentation. Very few studies have been reported regarding the toxicity of ionic liquids to bacteria.21 Furthermore, there is not any single report on the effects of ILs on probiotic bacteria like Propionibacteria.
Since ILs are or may some day be in widespread use, extensive research on the toxicological behavior of them is needed. Among various types of imidazolium-based ionic liquids, a vast majority of toxicity information is available for only a few kinds of anion types such as PF6, BF4, Br, Cl and MeSO4.21–24 However, to the best of our knowledge there is no information on the toxicity of imidazolium-based ILs with anions such as NO3, SCN, PTS and MeOSO3 on the microorganisms.
In this study, a wide series of imidazolium based ILs with various alkyl chain lengths and anion types were employed to investigate their toxicity on the probiotic strain of Propionibacterium freudenreichii as well as the effects of ILs on intracellular VB12 and extracellular PA biosynthesis. It is worth to say that, this is the first report on the effect of ILs on the production of an intracellular product of a bacterium. Toxicity study of ILs on probiotic bacteria, which are of interest in food, dairy and pharmaceutical industries may provide useful information for their usage not only in such industries, but also in their application in liquid–liquid extraction of biomaterials and extractive fermentation.
Results
Effect of ILs on the cell growth
The ionic liquids were added to the culture medium at the following concentrations: 2; 1; 0.5; 0.1 and 0.05% (w/v), except in the case of [OMIM][NO3], lower concentrations of 0.01 and 0.005% (w/v) were also used. Fig. 1A shows that 0.1% (w/v) and lower concentrations of [BMIM][BF4] had no effect on the growth of P. freudenreichii. At 0.5% (w/v) a short lag phase of 14 h was occurred. However, cell growth cannot be observed for concentrations higher than 0.5%. Only, in the case of 1% of the mentioned IL, a very long lag phase of about 96 h can be observed. Almost similar results were obtained by other researchers using the same IL.22,23 Ganske and Bornscheuer22 showed that [BMIM][BF4] was toxic at 1% (v/v) on the growth of Escherichia coli, Pichia pastoris and Bacillus cereus. Wang et al.23 also showed that Clostridium sp. can not grow in the presence of 1% (v/v) of this IL. [HMIM][BF4] showed different behavior. As seen in Fig. 1B, in the presence of 0.5% (w/v), no further growth was observed, while use of concentrations of the ionic liquid to 0.1 and 0.05% (w/v) led to a distinct delay in growth. For concentrations of 0.05 and 0.1% (w/v) of this IL, the lag phase of about 48 h and 72 h were occurred, respectively. | ||
| Fig. 1 Optical density during the growth of P. freudenreichii in the presence of different percents (w/v) of (A) [BMIM][BF4] and (B) [HMIM][BF4]. Data are mean values ± standard deviation of three replicates. Vertical bars on data points are ±SD of the mean (not shown when smaller than the symbols). | ||
[BMIM][PTS] had no negative effect on the growth in concentrations of 0.5% and lower. As can be observed in Fig. 2A, for this range of concentration, the growth of microorganism exceeds the control after 48 h. This indicated that [BMIM][PTS] has a positive effect on the growth in this area. Although, 0.5% (w/v) of this IL had no negative effect on the growth, doubling the concentration to 1% (w/v), substantially affected the growth (Fig. 2A) and led to a 72 h delay in growth. Higher concentrations completely inhibited the cell growth. P. freudenreichii tolerated 0.05% (w/v) of [HMIM][PTS] very well as indicated in Fig. 2B. Though higher concentrations delayed or inhibited the cell growth. In the case of 0.5% (w/v) of the IL, an interesting growth curve can be observed for the bacterium. Under such circumstances after a lag phase followed by an exponential phase, the bacterium reached to its death phase. In fact a complete growth curve was appeared.
 | ||
| Fig. 2 Optical density during the growth of P. freudenreichii in the presence of different percents (w/v) of (A) [BMIM][PTS] and (B) [HMIM][PTS]. Other details are as explained in Fig. 1. | ||
[BMIM][MeOSO3] and [HMIM][MeOSO3] showed similar results of cell growth at lower concentrations (Fig. 3A and B). Although at the concentration of 0.5% (w/v) of [HMIM][MeOSO3], a lag phase was observed, the final growth of P. freudenreichii has reached the growth of control. Both [BMIM][MeOSO3] and [HMIM][MeOSO3] inhibited the cell growth at the concentration of 1% and higher.
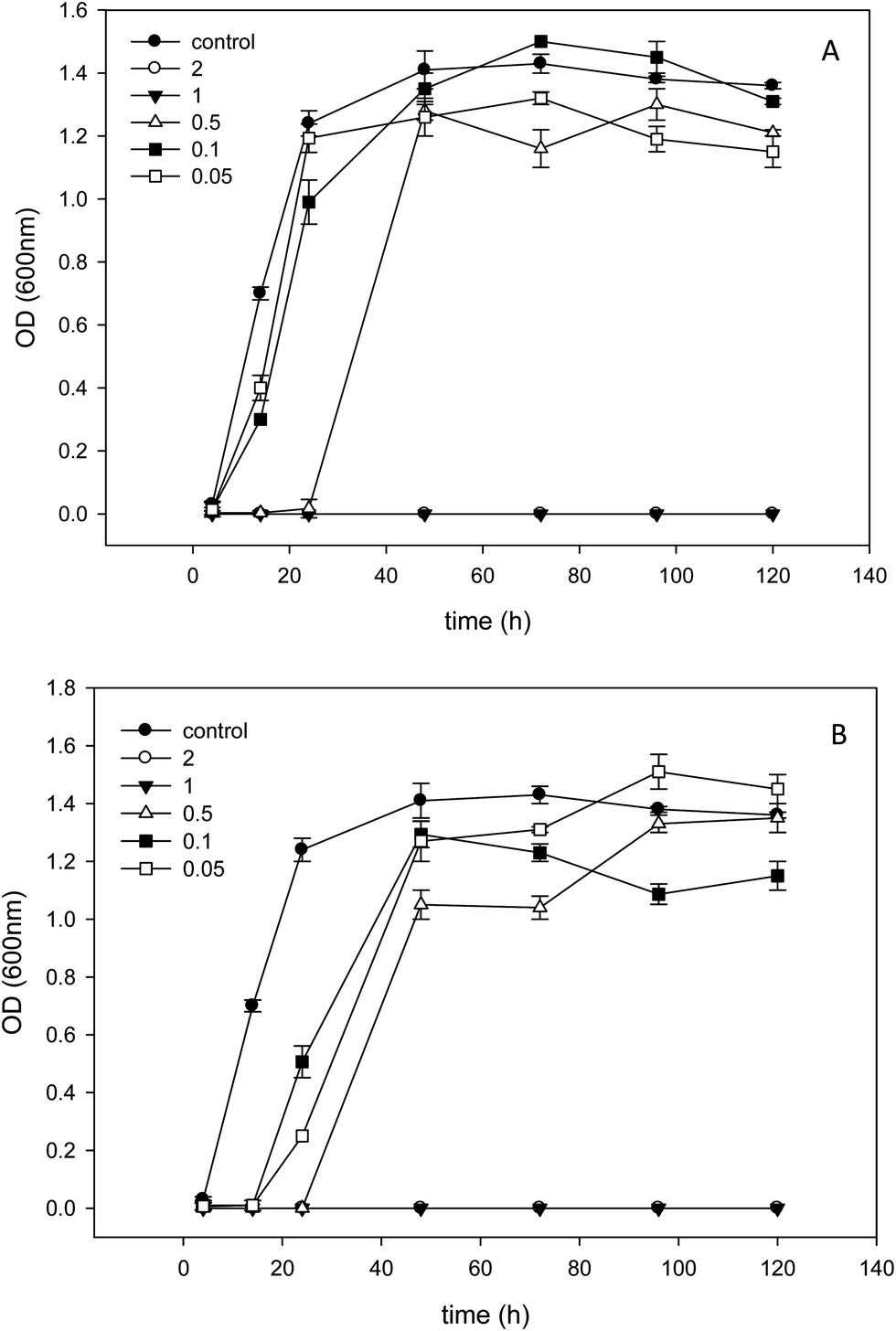 | ||
| Fig. 3 Optical density during the growth of P. freudenreichii in the presence of different percents (w/v) of (A) [BMIM][MeOSO3] and (B) [OMIM][MeOSO3]. Other details are as explained in Fig. 1. | ||
While P. freudenreichii could tolerate 0.5% (w/v) [BMIM][SCN] after a short delay, [OMIM][SCN] was inhibited cell growth at 0.5% and higher (Fig. 4A and B). The growth of bacterium in the presence of 0.1% [BMIM][SCN] was very similar to that of control, showing that this concentration has not positive nor negative effect on the growth. For [BMIM][SCN] at the concentration of 0.05%, growth was somewhat slower and the maximum growth starts later. While, the concentration of 0.5% of [BMIM][SCN] led to a 24 h lag phase, this amount completely inhibited cell growth using [OMIM][SCN].
 | ||
| Fig. 4 Optical density during the growth of P. freudenreichii in the presence of different percents (w/v) of (A) [BMIM][SCN] and (B) [OMIM][SCN]. Other details are as explained in Fig. 1. | ||
Interesting results were also obtained by ILs with the nitrate anion. Fig. 5 shows the effect of [BMIM][NO3], [HMIM][NO3] and [OMIM][NO3] on the cell growth of P. freudenreichii. Three alkyl chain lengths were employed. Concentrations below 0.5% of [BMIM][NO3] have not significantly influenced on the growth. 1% (w/v) of the IL led to long lag phase of 72 h. [HMIM][NO3] at 0.05 and 0.1% (w/v) didn't influence growth. However, 0.5% (w/v) led to lag phase and higher concentrations inhibited the cell growth completely. [OMIM][NO3] was the most toxic IL between all examined ILs. As seen in Fig. 5C, in the presence of 2, 1, 0.5, 0.1 and 0.05% (w/v) of this IL no growth was observed, hence, lower concentrations were examined. The concentration of 0.01% also inhibited the cell growth. P. freudenreichii starts to grow after 48 h lag phase in the presence of 0.005% of [OMIM][NO3]. Therefore, it can be concluded that this IL has a vigorous effect on the cell growth of P. freudenreichii.
 | ||
| Fig. 5 Optical density during the growth of P. freudenreichii in the presence of different percents (w/v) of (A) [BMIM][NO3], (B) [HMIM][NO3] and (C) [OMIM][NO3]. Other details are as explained in Fig. 1. | ||
Using the data from Fig. 1–5, the range of toxic effect of ILs in term of IC50-48h can be obtained. The amounts of IC50-48h have been summarized in Table 1. As it is clear from the definition of this parameter, the lower the amount of IC50-48h, the more the toxicity of the IL. Hence, the most toxic IL among all is [OMIM][NO3] with IC50-48h equal to 0.005% (w/v). [HMIM][BF4] with IC50-48h equal to 0.05% (w/v) is in the second position in term of toxicity. As stated above, almost similar toxicity can be observed between [BMIM][MeOSO3] and [OMIM][MeOSO3] with IC50-48h between 0.5 and 1.
h)
| Ionic liquid | IC50-48h |
Ionic liquid | IC50-48h |
|---|---|---|---|
| [BMIM][BF4] | 0.5–1 | [BDMIM][SCN] | 0.5–1 |
| [HMIM][BF4] | 0.05 | [OMIM][SCN] | 0.1–0.5 |
| [BMIM][PTS] | 1 | [BMIM][NO3] | 1 |
| [HMIM][PTS] | 0.5 | [HMIM][NO3] | 0.5 |
| [BMIM][MeOSO3] | 0.5–1 | [OMIM][NO3] | 0.005 |
| [OMIM][MeOSO3] | 0.5–1 |
Effect of ILs on the end products formation
In order to determine how ILs can affect the production of intracellular and extracellular bioproducts, the formation of intracellular (i.e. VB12) and extracellular (i.e. PA) products of P. freudenreichii were investigated.Table 2 shows the amount of VB12 and PA produced by P. freudenreichii after 120 h of fermentation time. The amounts of produced bioproducts for various concentrations of ILs are presented in this table. In order to compare these data better, the bar diagrams of them can be seen as Fig. S1–S5 of the ESI.†
| Ionic liquid | Intracellular VB12 (mg l−1) | Extracellular PA (g l−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2% | 1% | 0.50% | 0.10% | 0.05% | 0.01% | 2% | 1% | 0.50% | 0.10% | 0.05% | 0.01% | |
| a PA in control: 12.41 g l−1, VB12 in control: 2.23 mg l−1, N.D: not determined, all data groups are significant with P-value < 0.05. | ||||||||||||
| [BMIM][BF4] | 0 | 0 | 0.66 | 1.34 | 1.2 | N.D | 0 | 2.2 | 12.1 | 12 | 12.33 | N.D |
| [HMIM][BF4] | 0 | 0 | 0 | 0.41 | 0.91 | N.D | 0 | 0 | 0 | 4.1 | 8.7 | N.D |
| [BMIM][PTS] | 0 | 0.67 | 1.22 | 1.82 | 1.63 | N.D | 0 | 0.5 | 12.9 | 13.1 | 13.1 | N.D |
| [HMIM][PTS] | 0 | 0 | 0.55 | 1.39 | 1.94 | N.D | 0 | 0.1 | 8 | 11.1 | 12 | N.D |
| [BMIM][MeOSO3] | 0 | 0 | 1.14 | 1.18 | 1.1 | N.D | 0 | 0.1 | 9.7 | 10.5 | 11.8 | N.D |
| [OMIM][MeOSO3] | 0 | 0 | 1.22 | 2.1 | 1.1 | N.D | 0 | 0.7 | 8.3 | 9.7 | 12.1 | N.D |
| [BMIM][SCN] | 0 | 0 | 1 | 1.4 | 0.74 | N.D | 0 | 0.5 | 6.9 | 9.6 | 9.6 | N.D |
| [OMIM][SCN] | 0 | 0 | 0 | 1.9 | 2.1 | N.D | 0 | 0.5 | 1.1 | 8.6 | 8.9 | N.D |
| [BMIM][NO3] | 0 | 0.34 | 1.55 | 2.18 | 1.5 | N.D | 0 | 2 | 4.5 | 8 | 8.1 | N.D |
| [HMIM][NO3] | 0 | 0 | 0 | 1.36 | 0.94 | N.D | 0 | 0 | 0.9 | 8.1 | 8.05 | N.D |
| [OMIM][NO3] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.4 | 0.5 | 2.9 |
The amount of produced PA is directly related to the concentration of IL almost in all cases, i.e. increasing the IL percentage reducing the PA amount, with the increase of IL percent, the amount of PA has been reduced. On the other hand, according to Fig. 1–5, the growth of bacterium relates to the ILs' concentration. Therefore, it can be concluded that the PA production is also growth related. Maximum production of PA was achieved in the presence of [BMIM][PTS] (13.1 g l−1) which was more than the control (12.41 g l−1). ILs with NO3 anion produced lower amounts of PA compared to other ILs. Among them [OMIM][NO3] produced the least amount of PA. This is because of very weak growth of P. freudenreichii in the presence of this IL.
While the PA production was related to the concentration of ILs, VB12 production showed very different behavior. The relation between IL concentration and VB12 production was almost complex. Surprisingly, all ILs with butyl chain induced the cells to produce VB12 in higher amounts at the concentration of 1% (w/v) than at 0.05% (w/v). It means that increasing in IL concentration, stimulates the cells to produce higher amounts of intracellular product. However, the amount of VB12 reduced at the IL concentration of 0.5% in most cases. In the case of [BMIM][MeOSO3], the amount of produced VB12 at the presence of 0.5% IL is even more than for 0.05%. This showed that increasing of IL concentration 10 times have resulted in the increase of VB12 from 1.1 to 1.14 mg l−1. However, among ILs with longer alkyl chain, only [OMIM][MeOSO3], showed this behavior. In the case of other ILs, the amount of produced VB12 was directly related to the concentration of ILs.
Discussion
Complexity of chemical nature of ILs has made it difficult to decide on their real cause of toxicity on microorganisms. However, alkyl chain length, the various functional groups, cationic head groups, and types of anions are active variables that are assumed to influence the toxicity of these compounds. Stolte et al.25 showed that the cationic head group has a minor effect on the ILs' toxicity while, alkyl chain length and type of anions have the main effect.Effect of alkyl chain length on toxicity
In order to explain the well-known effect of IL alkyl chain length on toxicity towards the probiotic microorganism under investigation, the cell growth diagrams of Fig. 1–5 as well as Tables 1 and 2 should be considered. All of the small alkyl chain length ILs, which are tested have a butyl chain (Table 3), the other ILs have hexyl or octyl chains. Therefore, considering the ILs with similar anions, the chain length effect can be compared. In all cases, it can be concluded that the longer the chain length of cation, the more toxic the IL toward microorganism in terms of cell growth and IC50-48h. For example, as can be seen in Fig. 5, IL with nitrate anion and three different alkyl chains of C4, C6 and C8 can explain this effect. While the microorganism can tolerate 0.5% (w/v) of the IL with C4 chain, it doesn't have the ability to sustain similar concentration using the IL with C6 chain. Furthermore, the microorganism can not tolerate very lower concentrations of 0.01 and 0.005% (w/v) of IL with C8 chain. This can also be observed in Table 1, in which the IC50-48h for the IL with nitrate anion and three different alkyl chains of C4, C6 and C8 are 1, 0.5 and 0.005, respectively. This trend is maintained with the other ILs, with the similar anions exhibiting a far higher sensitivity to alkyl chain length. This observation has been demonstrated by many researchers.17,24 Latala et al.24 showed that alkyl chain length has a direct relation with the degree of toxicity toward green algae.
| Ionic liquid | Chemical structure | Acronym | Viscosity (Pa s) | Density (g cm−3) | Molecular weight |
|---|---|---|---|---|---|
| a No data available.b Solid form. | |||||
| 1-Butyl-3-methylimidazolium tetrafluoroborate |  |
[BMIM][BF4] | 233 (25 °C) | 1.12 (25 °C) | 226.02 |
| 1-Butyl-3-methylimidazolium tosylate |  |
[BMIM][PTS] | a | a | 310.41 |
| 1-Butyl-3-methylimidazolium nitrate |  |
[BMIM][NO3] | 266 (20 °C) | 1.15 (20 °C) | 201.22 |
| 1-Butyl-3-methylimidazolium thiocyanate |  |
[BMIM][SCN] | a | 1.02 (20 °C) | 197.30 |
| 1-Butyl-3-methylimidazolium methyl sulfate |  |
[BMIM][MeOSO3] | a | 1.21 (20 °C) | 250.32 |
| 1-Hexyl-3-methylimidazolium tetrafluoroborate |  |
[HMIM][BF4] | 314 (20 °C) | 1.14 (20 °C) | 254.08 |
| 1-Hexyl-3-methylimidazolium tosylate |  |
[HMIM][PTS] | b | b | 338.41 |
| 1-Hexyl-3-methylimidazolium nitrate |  |
[HMIM][NO3] | 804 (20 °C) | 1.19 (20 °C) | 229.22 |
| 1-Octyl-3-methylimidazolium methylsulfate |  |
[OMIM][MeOSO3] | a | a | 306.42 |
| 1-Octyl-3-methylimidazolium thiocyanate |  |
[OMIM][SCN] | a | a | 239.3 |
| 1-Octyl-3-methylimidazolium nitrate |  |
[OMIM][NO3] | 1238 (20 °C) | a | 243.22 |
Although increasing in the alkyl chain length, reduced the production of PA (Table 2) almost in all cases, its effect on the production of intracellular VB12 is not clear. In some cases by increasing the alkyl chain length, the production of VB12 increased and in some others decreased. This showed that some ILs have a positive effect on intracellular VB12 biosynthesis in a certain concentrations.
The underlying mechanism responsible for the real cause of toxicity remains unknown. However, in order to explain the reason of toxicity of ILs to bacteria, it is necessary to take a look at their structure in one hand, and considering the interaction of ILs with the cell membrane on the other hand. It is known that the structure of many ILs is similar to surfactants and/or biosurfactants.26 Surfactants and biosurfactants have a large head group with a long hydrocarbon chain. It is proved today that these compounds have antimicrobial activity and inhibit the growth of microorganisms. Besides, Stolte et al.25 showed that increasing IL chain length would lead to the increase in lipophilicity. Therefore, ILs containing longer alkyl chains exhibit an adsorption potential through simultaneous hydrophobic and ionic interactions (in the same way as surfactants), they have a greater ability to accumulate at the bacterial membrane–water interface, which causes the disruption of the integral membrane.27
While the disruption mechanism remains unclear, this may be related to the type of cell membrane. Disruption of cell membrane for Gram positive and Gram negative bacteria may have different mechanisms. The outer membrane of Gram-negative bacteria is composed of lipopolysaccharides (LPS), phospholipids and lipoproteins, covalently linked to the peptidoglycan layer through hydrophobic interactions.28 Therefore, ILs can interact with this hydrophobe sites and disrupt synthetic membranes.29 Sotirova et al.30 investigated the impact of biosurfactants (which have similar structures to ILs) on the cell properties of a Gram negative bacterium. They showed that in the presence of biosurfactant the amount of LPS in the cell membrane reduced to 78% lower than control. In similar manner, ILs can interact with the cell membrane, as well.
For Gram positive bacteria, which have not lipopolysaccharides in their membrane, the outer layer of cell membrane is composed of a thick peptidoglycan layer. As Murugesan et al.31 showed, some ILs could dissolve glycosaminoglycans, which its structure is very similar to peptidoglycan. Hence, it can be assumed that the IL interact with the outer layer of the membrane of Gram positive bacteria and dissolve the peptidoglycan layer. In this way, the cell wall will be interrupted.
Effect of anion type on toxicity
It has been demonstrated that IL toxicity is affected by the choice of anion.17,24,25 In the present case, the anion effect can be easily examined by comparing ILs with various anions of BF4, PTS, NO3, MeOSO3, SCN according to Fig. 1–5 and Tables 1 and 2. Generally, P. freudenreichii was very sensitive to the changes in anion. NO3 was the most toxic anion among all ILs. This is clear by comparing the ILs with similar alkyl chain length and different anions. PTS however was the least toxic one. In order to explain the reason of toxicity of ILs with BF4 anions, it had been postulated that BF4 could undergo hydrolysis. This would lead to the formation of fluoride and the consequent potential of the toxic effect.32 This was later confirmed by other literatures.25,33According to Table 2, among ILs with different anions, [HMIM][BF4] strongly inhibited the production of both intracellular and extracellular products, even more than [HMIM][NO3]. However, it seems that alkyl chain effect is predominant to anion effect. This is clear from Table 2 that increase in alkyl chain length has a more vigorous effect on the end products formation than changing anion type of the molecule.
Conclusion
Effects of various kinds of ionic liquids on the cell growth and end products formation of P. freudenreichii were investigated for the first time. All eleven kinds of ILs affected P. freudenreichii growth and end products formation. However, each IL affected differently. [OMIM][NO3] inhibited the growth and production vigorously. On the other hand, ILs with PTS anion were less toxic compared to other ILs. While the production of PA was related to the concentration of IL, a complex relation for intracellular product was observed. Small chain length ILs improved the production of VB12 in some extent, while large chain length ILs generally had negative effect on the production.Materials and method
Chemicals, ionic liquids and microorganisms
All chemicals were obtained from Merck (Germany) unless otherwise stated. ILs which have been used in this work are presented in Table 3. All ILs are synthesized in our laboratory according to our previous works.1,2,34–36 ILs are imidazolium based with different anions and different alkyl chain lengths. Therefore, the comparison between ILs, can be assessed at anion type and alkyl chain length. Propionibacterium freudenreichii subsp. freudenreichii PTCC 1674 was purchased from Persian Type Culture Collection in lyophilized state and stored at −70 °C in glycerol stocks. This strain is Gram-positive, non-spore forming, rod-shape, and aerotolerant anaerobic bacterium.Growth and cultivation
For inoculum preparation, the lyophilized bacterium from glycerol stock (1.5 ml) was thawed and precultured in nutrient broth medium at 120 rpm, 30 °C for 18 h. 2% (v/v) inoculation was performed from this culture to the production medium containing (g per liter of distilled water): glucose 30, peptone 5, yeast extract 10, KH2PO4 1, K2HPO4 1, (NH4)2NO3 4, MgCl2·6H2O 1, MnCl2·2H2O 0.002 and betaine 5, 5,6-dimethylbenzimidazole (DMB) 0.005 CoCl2·6H2O 0.02, FeSO4·7H2O 0.005 and incubated on a temperature controlled Kuhner shaker incubator (Germany) at 130 rpm and 30 °C. pH before autoclaving adjusted between 6.8 and 7 using 3 N NaOH.Extraction and analysis
The concentration of VB12 was determined by high performance liquid chromatography (HPLC) reported elsewhere.37 Briefly, the culture broth was centrifuged (7000 × g, 15 min) to harvest cells. The pellet was washed with 0.2 M phosphate buffer (pH 5.5), centrifuged again (7000 × g, 15 min) and the cells were dissolved in 0.1% KCN (pH 6.0) and then autoclaved at 121 °C for 15 min. After cold, the sample was centrifuged to remove solid. The supernatant was filtered (0.45 μm nylon filters) and analyzed with HPLC. HPLC was performed on a reversed-phase C18 column (Shimadzu 250 mm × 4.6 mm; 5 μm particle size), UV detection at 361 nm, flow rate of 1 ml min−1, oven temperature at 25 °C, and injection volume of 20 μl. Cyanocobalamin (VB12) standards (α-(5,6-dimethylbenzimidazolyl)cyanocobamid) purchased from Da Jung CO. (South Korea) at concentrations ranging from 0.5 to 20 μg ml−1 were used for calibration. Each sample was injected twice and means were reported.Propionic acid concentration in the broth was obtained using the same HPLC system, with the mobile phase of the solution of 1 mmol l−1 sulfuric acid plus 8 mmol l−1 Na2SO4 in deionized water (pH 2.8). Flow rate was set to 1 ml min−1, oven temperature at 25 °C, and injection volume was 20 μl. Propionic acid standards at concentrations ranging from 1 to 30 g l−1 were used for calibration.
Toxicity of ionic liquids
IL toxicity is usually determined by different criteria such as inhibition of cell growth, cell viability or even inhibition of the metabolism measured through product or substrate uptake. To homologate the toxicity data, it is conventional to define the inhibitory concentration (IC50) at which cell growth is inhibited at 50% relative to a control deprived of IL.23 Hence, in this study, besides the evaluation of bacterial growth over time, we also quantified the toxicity of each IL by determining the IC50-48h; this notation represents the concentration of ILs that induces 50% of the maximum inhibitory response after 48 h exposure. To evaluate the inhibition of cell growth in the presence of ILs, stock solutions of ILs were prepared in deionized water and deoxygenated by purging with N2 gas. ILs were sterile filtered through a 0.22 μm filter and then aseptically added to 50 ml of mineral-salts (MS) medium in 250 ml flasks. Cultures containing MS medium with and without ILs were inoculated with 2% P. freudenreichii. In specific time intervals, 500 μl sample was withdrawn aseptically and the growth of bacterium was determined by measuring optical density (OD) at 600 nm using a UV-visible spectrophotometer (Perkin-Elmer, model Lambda25, USA).
In order to investigate the effect of ILs on the end products formation, the production of two essential bioproducts of P. freudenreichii i.e. intracellular VB12 and extracellular PA were studied. Production of VB12 and PA after 120 h was considered as a result (Fig. S6†). All data are the average of triplicate experiments.
Statistical analysis
All data were analyzed using the SPSS 11.5 statistical analysis system. A one-way analysis of variance was used to determine whether a significant difference existed between the treated groups and controls. Data were expressed as mean ± standard deviation (SD) and differences were considered statistically significant if P < 0.05.References
- B. Mokhtarani, A. Sharifi, H. Mortaheb, M. Mirzaei, M. Mafi and F. Sadeghian, J. Chem. Thermodyn., 2009, 41, 1432–1438 CrossRef CAS PubMed.
- B. Mokhtarani, A. Sharifi, H. Mortaheb, M. Mirzaei, M. Mafi and F. Sadeghian, J. Chem. Thermodyn., 2009, 41, 323–329 CrossRef CAS PubMed.
- J. M. Ma, L. L. Cai, B. J. Zhang, L. W. Hu, X. Y. Li and J. J. Wang, Ecotoxicol. Environ. Saf., 2010, 73, 1465–1469 CrossRef CAS PubMed.
- T. Welton, Coord. Chem. Rev., 2004, 248, 2459–2477 CrossRef CAS PubMed.
- M. Koel, Crit. Rev. Anal. Chem., 2005, 35, 177–192 CrossRef CAS.
- V. A. Cocalia, K. E. Gutowski and R. D. Rogers, Coord. Chem. Rev., 2005, 250, 755–764 CrossRef PubMed.
- P. Kubisa, J. Polym. Sci., Part A: Polym. Chem., 2005, 43, 4675–4683 CrossRef CAS.
- H. Xue, R. Verma and J. M. Shreeve, J. Fluorine Chem., 2006, 127, 159–176 CrossRef CAS PubMed.
- M. Antonietti, D. Kuang, B. Smarsly and Y. Zhou, Angew. Chem., Int. Ed., 2004, 43, 4988–4992 CrossRef CAS PubMed.
- R. J. Bernot, M. A. Brueseke, M. A. Evans-White and G. A. Lamberti, Environ. Toxicol. Chem., 2005, 24, 87–92 CrossRef CAS.
- K. M. Docherty and C. F. Kulpa, Green Chem., 2005, 7, 185–189 RSC.
- Z. Yang and W. Pan, Enzyme Microb. Technol., 2005, 37, 19–28 CrossRef CAS PubMed.
- M. Matsumoto, K. Mochiduki, K. Fukunishi and K. Kondo, Sep. Purif. Technol., 2004, 40, 97–101 CrossRef CAS PubMed.
- Y. Kim, Y. Choi, J. Park, S. Lee, Y. Yang, H. J. Kim, T. Park, Y. H. Kim and S. H. Lee, Bioresour. Technol., 2012, 109, 312–315 CrossRef CAS PubMed.
- J. Vitz, T. Erdmenger, C. Haensch and U. S. Schubert, Green Chem., 2009, 11, 417–424 RSC.
- C. Liu, F. Wang, A. R. Stiles and C. Guo, Appl. Energy, 2012, 92, 406–414 CrossRef CAS PubMed.
- Y. V. Nancharaiah and A. J. Francis, Bioresour. Technol., 2011, 102, 6573–6578 CrossRef PubMed.
- Y. Piao, N. Kawaraichi, R. Asegawa, P. Kiatpan, H. Ono, M. Yamashita and Y. Murooka, J. Biosci. Bioeng., 2004, 97(5), 310–316 CrossRef CAS.
- K. Miyano, K. Ye and K. Shimizu, Biochem. Eng. J., 2000, 6, 207–214 CrossRef CAS.
- Z. Gu, L. Bonita, A. Glatz and C. E. Glatz, Biotechnol. Bioeng., 1998, 57, 454–461 CrossRef CAS.
- M. Matsumoto, K. Mochiduki and K. Kondo, J. Biosci. Bioeng., 2004, 98(5), 344–347 CrossRef CAS.
- F. Ganske and U. T. Bornscheuer, Biotechnol. Lett., 2006, 28, 465–469 CrossRef CAS PubMed.
- H. Wang, S. V. Malhotra and A. J. Francis, Chemosphere, 2011, 82, 1597–1603 CrossRef CAS PubMed.
- A. Latala, M. Nedzi and P. Stepnowski, Green Chem., 2009, 11, 580–588 RSC.
- S. Stolte, M. Matzke, J. Arning, A. Boschen, W. Pitner, U. Welz-Biermann, B. Jastorff and J. Ranke, Green Chem., 2007, 9, 1170–1179 RSC.
- P. J. Scammells, J. L. Scott and R. D. Singer, Aust. J. Chem., 2005, 58, 155–169 CrossRef CAS.
- W. Mrozik, C. Jungnickel, M. Skup, P. Urbaszek and P. Stepnowski, Environ. Chem., 2008, 5, 299–306 CrossRef CAS.
- J. Sikkema, J. A. de Bont and B. Poolman, Microbiol. Rev., 1995, 59, 201–222 CAS.
- K. O. Evans, Colloids Surf., A, 2006, 274, 11–17 CrossRef CAS PubMed.
- A. Sotirova, D. Spasova, E. Vasileva-Tonkova and D. Galabova, Microbiol. Res., 2009, 164, 297–303 CrossRef CAS PubMed.
- S. Murugesan, J. M. Wiencek, R. X. Ren and R. J. Linhardt, Carbohydr. Polym., 2006, 63, 268–271 CrossRef CAS PubMed.
- P. Stepnowski, A. C. Składanowski, A. Ludwiczak and E. Laczynska, Hum. Exp. Toxicol., 2004, 23, 513–517 CAS.
- M. Matzke, S. Stolte, A. Boschen and J. Filser, Green Chem., 2008, 10, 784–792 RSC.
- B. Mokhtarani, L. Valialahi, K. T. Heidar, H. Mortaheb, A. Sharifi and M. Mirzaei, Fluid Phase Equilib., 2012, 334, 65–69 CrossRef CAS PubMed.
- B. Mokhtarani, J. Musavi, M. Parvini, M. Mafi, A. Sharifi and M. Mirzaei, Fluid Phase Equilib., 2013, 341, 35–41 CrossRef CAS PubMed.
- B. Mokhtarani, A. Sharifi, H. Mortaheb, M. Mirzaei, M. Mafi and F. Sadeghian, J. Chem. Eng. Data, 2010, 55, 3901–3908 CrossRef CAS.
- S. Hugenschmidt, S. M. Schwenninger, N. Gnehm and C. Lacroix, Int. Dairy J., 2010, 20, 852–857 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra00925h |
| This journal is © The Royal Society of Chemistry 2014 |