Augmented biocontrol action of silica nanoparticles and Pseudomonas fluorescens bioformulant in maize (Zea mays L.)
Suriyaprabha Rangaraja,
Karunakaran Gopalua,
Prabhu Muthusamya,
Yuvakkumar Rathinama,
Rajendran Venkatachalam*a and
Kannan Narayanasamyb
aCentre for Nanoscience and Technology, K. S. Rangasamy College of Technology, Tiruchengode 637215, Tamil Nadu, India. E-mail: veerajendran@gmail.com; Fax: +91-4288-27488; Tel: +91-4288-274741-4
bDepartment of Biotechnology, K. S. Rangasamy College of Arts and Science, Tiruchengode 637215, Tamil Nadu, India
First published on 15th January 2014
Abstract
Nanosilica and Pseudomonas sp. was treated in soil to enhance the biocontrol activity against pathogens in maize. The leaf hardness shows an increase in surface roughness and elevated phenols while treating nanobiocomposites. Higher phenols and hardness induce silica accumulation in leaves and initiate more effective physical barriers for crop protection.
Protection and nutrition strategies for plants and the environment are periodically attempted to provide new solutions to existing problems such as drought, climate change and soil nutrient deficiency. To tackle this, crop yield improvement by developing disease resistance or stress tolerant plants is one of the important approaches.
Nanomaterials usage is currently widening in agriculture through different applications such as pesticides, fertilizers, and nanonutrients.1–3 The interactions between crops and nanomaterials lead to different roles in agricultural biotechnology. Recent studies show that few metal oxide nanoparticles and carbon-based materials promote plant growth because of their smaller size.4,5
Because of the unique properties of nanomaterials, one can harness innovative approaches of using nanotechnology in agriculture.6,7 An anticipated contribution of nanoparticles is to stabilise biocontrol preparations, thereby reducing the environmental hazard,6 which is the current area of interest. Silicon or silica is considered as an essential nutrient “anomaly” for monocotyledons. Several reports are available on the role of silicon and silica sources in plants and their extraction processes.8–10 However, the efficiency for soil application relies on the type and expense of silica source used. Amorphous nanosilica shows promising potential as a biopesticide.1,11 In fact, an enhanced SiO2 accumulation in nanosilica treated maize is evident from our previous study.2 Primarily, the effect of silica nanoparticles (SNPs) on conferring mechanical protection in monocotyledons is yet to be explored for disease resistance as exogenous application is insufficient for broad biocontrol action.
Pseudomonas fluorescens is a known biocontrol agent that protects a range of crop plants from root pathogens.12,13 However, their delivery and biocontrol activity require a potential carrier for soil or foliar applications. Some of the constraints in widespread usage of microorganisms are short shelf life, lack of carrier materials, and transportation and storage. In addition, artificially introduced plant growth promoting rhizobacteria (PGPR) can initially colonize roots at 107–108 Colony Forming Units (CFU) g−1, but these levels decline in a few weeks.14 Moreover, the interaction of nanoparticles with bacteria and their influence on soil communities and plants are evident from previous studies that nanoparticle can penetrate into microorganism via cell wall pores and accumulate/react with cell bodies.15,16 Alternatives to agrochemicals are bioformulations which gain acceptance in terms of their applicability, efficiency and eco-friendly nature, though their on-field stability and shelf life are the major concerns. On the basis of the concepts related to the interactions between bacteria and nanoparticles,15–17 an approach on the application of biocontrol agent and SNPs to soil is discussed in the present study.
The work on combined effort of applying nanosilica with a biocontrol agent is found to be scanty. Therefore, in this study, an emphasis is laid on the interaction mechanisms of SNPs with an extensively studied group of fluorescent pseudomonads in maize for assuming better biocontrol action. This unique approach is aimed to direct the path to improve the defense action via nanosilica-mediated physical strength exerted to monocotyledons.
In this study, analytical grade (AR) chemicals were used as received from Merck (Mumbai, India) without any further purification. Conventional microsilica (SiO2) (MW 60.08; 40–150 mesh; 99.99%) and prepared amorphous nanosilica (20–40 nm; 99.72%) extracted from natural plant biomass rice husk2 were used as silica sources.
The fine sandy loam soil was collected from the agriculture land at Tiruchengode (Tamil Nadu, India) for pot studies. The experiment was set in a split-plot design with SNPs as the main plot factor. The soil was mixed with nanosilica at different concentrations, namely 5, 10 and 15 kg ha−1 (hereafter termed as N5, N10, and N15) and with bulk silica at 15 kg ha−1 (hereafter termed as B15). The above soil mixtures were further used for the treatment of biocontrol agent, Pseudomonas fluorescens. The bacterial culture (NCIM 2390) is collected from National Chemical Laboratory (NCL), National Collection of Industrial Microorganisms, Pune India. A loopful of stock fluorescent pseudomonad cultures was inoculated and grown in Erlenmeyer flasks (250 ml) containing 100 ml nutrient broth for 24 h on a rotary shaker (100 rpm) at 28 ± 2 °C. Supernatant was removed after centrifugation at 8000 rpm for 10 min at 4 °C and then, the pellet was diluted with adequate amount of sterile distilled water to obtain a bacterial suspension of 108 CFU ml−1. The culture was mixed aseptically with the soils amended with silica particles. Maize hybrid seeds (Zea mays L.; TIP TOP) purchased from Rasi Hybrid Seeds (Attur, Tamil Nadu, India) were sown in pots after 24 h of seed incubation. The maize grown in the soil without silica sources and biocontrol agent was kept as a control.
During 10 days of maize growth, roots were grown enough to collect soil from rhizosphere. The rhizosphere soil supplemented with silica sources and biocontrol agent was collected from the pots and was subjected to serial dilution followed by spread plate technique. The total population of P. fluorescens was enumerated from the soil using King's B selective media. Cultures of each triplicate samples were incubated at 28 ± 2 °C for 48 h. Growth of discrete colonies were determined and the population was expressed as CFU.
Significant differences in the bacterial population, phenol and enzyme activity with the regimes of silica particles in maize were analyzed by one way analysis of variance. The mean values of triplicate samples were compared by Duncan and Tukey's test using Statistical Package for Social Sciences software (version 16.0; SPSS, IL, USA) at 5% significant level (p < 0.05).
Essential resistance mechanism in plants other than defense compounds is the physical strength. The degree of physical strength attained in leaves by SNPs can be elucidated from the leaf hardness tests using nanomechanical testing systems.† Fig. 1 shows the topography of SNPs-treated maize leaves for the applied force (μN). The observed color contrast in the images shows that the roughness exists on leaf surface at a series of scanning displacement (nm).
 | ||
| Fig. 1 Surface topography of maize leaf surface. (a) Control (b) bulk silica and (c) nano silica. | ||
A thick section of leaves in nanosilica-treated maize is observed i.e., roughness and hardness (Table 1) are found to be respectively, 1640 nm and 26.46 MPa (Fig. 1c). In contrast, bulk and control leaves possess considerably less roughness and hardness value than nanosilica treated samples. The displacement of piezoelectric tip as a function of applied force on the leaf surface is graphically represented in Fig. 2. The depth curve is gradually decreased with an increase in force from nano, bulk and control leaves of maize which is clearly seen in the graph.
| Samples | Mean height (nm) | Roughness (nm) | Hardness (MPa) |
|---|---|---|---|
| Control | −0.001 | 368.0 | 1.51 |
| Bulk silica | −0.03 | 383.4 | 3.25 |
| Nano silica | 1.4 | 1640 | 26.46 |
 | ||
| Fig. 2 Mechanical indentation curve with respect to applied force on the leaf surface. | ||
Generally, SNPs are physiosorbed by the cuticular lipids, disrupting the protective barrier and thereby causing death of insects purely by physical means.12 Moreover, the surface charge-modified hydrophobic SNPs (3–5 nm) are successfully used to control a range of agricultural insects and animal ecto-parasites of veterinary importance.18 Further, SNPs are also applied as thin films on seeds to decrease fungal growth and boost cereal germination.19,20
Pseudomonas population is enumerated after 48 h of incubation in King's B medium and is illustrated in Fig. 3. Higher population (4.4 × 104 CFU g−1) is induced at N15 when compared to control (2.6 × 104 CFU g−1) and bulk (3.2 × 104 CFU g−1) samples. The efficiency of nanosilica in improving the soil biomass due to the treatment with silica and biocontrol agent is evident in this observation.
 | ||
| Fig. 3 Total population of pseudomonads in soil treated with silica sources. | ||
In our previous study, enhanced population of beneficial bacteria is noticed in the sandy loam soil treated with TiO2 and ZrO2 nanoparticles wherein a higher concentration led a decrease in population.21 On the contrary, our results show that an increase in nanosilica concentration increases the bacterial population. Therefore, it is inferred that if nanosilica enhances the bacterial population, it promotes the degree of biocontrol action against maize pathogen in soil.
After ten days of maize growth, the maize roots sample were collected and estimated for polyphenol oxidase (PPO), peroxidase phenylalanine ammonia lyase activity and phenol content.‡ The observed PPO activity in Fig. 4 indicates an increase in solution intensity with an increase in the incubation period after adding the substrate to the root extract. However, a decrease in enzyme content in the roots is noticed with an increase in the concentration of nanosilica treatment, particularly at N15. On the other hand, the control and bulk silica treated maize roots do not show any effect on enzyme content.
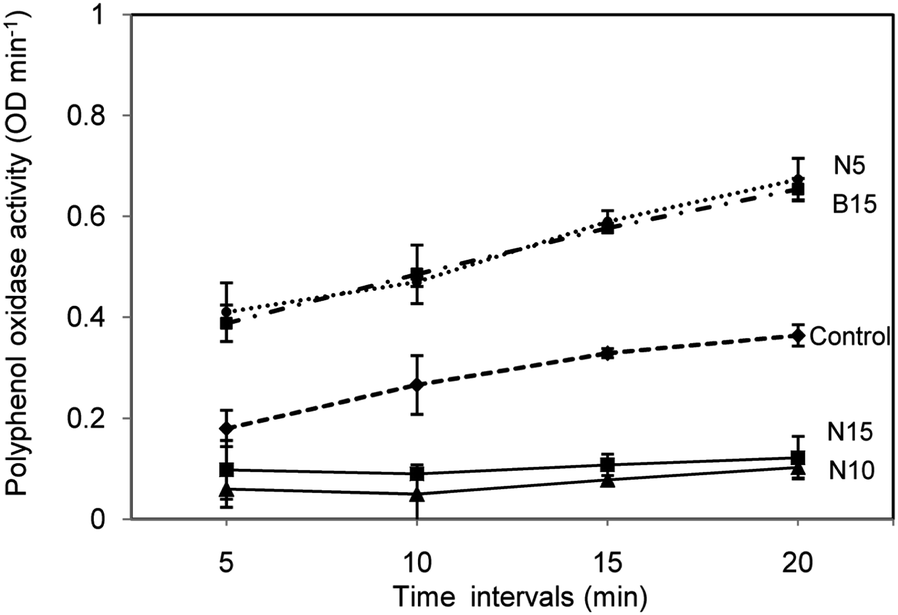 | ||
| Fig. 4 Estimation of PPO activity in maize roots treated with silica sources and Pseudomonas at different time intervals. | ||
The suppressed enzyme expression may indirectly convey lesser stress in maize. In addition, peroxidase and PAL activity of maize roots treated with Pseudomonas and different regimes of nanosilica are shown in Fig. 5 and 6, respectively.
 | ||
| Fig. 5 Estimation of Peroxidase activity in roots treated with silica sources and Pseudomonas at different time intervals. | ||
 | ||
| Fig. 6 PAL activity in roots treated with silica sources and Pseudomonas. | ||
On comparison of all treatments, N10 and N15 show reduced expression of peroxidases at a decrease in time interval with an increase in concentration. However, less enzyme activity is observed in sample B15 than N5 and control. The above results are in correlation with those of earlier studies that bulk silicon treatment induces damping-off mechanisms among defense compounds.16,22 Moreover, the application of silica is being instrumental in the formation of such defense compounds through the alteration of gene expression and afford defense against biotic stresses.23,24 Thus, it concludes an improved biocontrol action in maize crops while treating nanoscale silica. Hence, silicon-mediated defense mechanisms in monocotyledons support the application of SNPs formulated with P. fluorescens to achieve a broad spectrum of biocontrol activity.
Total phenol content in a methanolic extract of maize roots is observed to be increased in N15 (4.88 μg ml−1) among all treatments (Fig. 7). However, in B15 (2.17 μg ml−1) and control (2.46 μg ml−1), it is not differed at 5% significant level (p < 0.05).
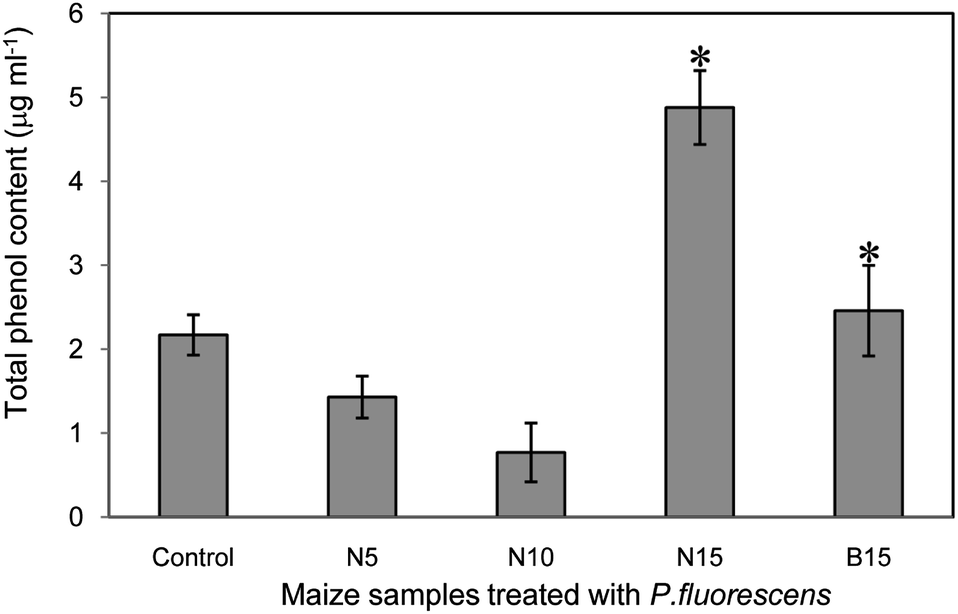 | ||
| Fig. 7 Total phenol content in maize roots treated with silica sources and Pseudomonas. * indicate the significant difference obtained by Duncans' test at 5% level (p < 0.05). | ||
A sharp rise in phenolic content at N15 in combination with biocontrol agent is observed even after three trials, which emphasises the induced resistance. These elevated levels of phenol may be due to enhancement of fungal resistance compounds released by P. fluorescens. Although many phenolic compounds in plant may not have biocontrol activity per se, the oxidation products of pre-existing phenols and bacterial extracellular enzymes might have biocontrol action.16 This is also due to the fact that soluble silica can easily forms complex with phenolic compounds, thereby strengthening the mechanical resistance of plant cells.24 A similar phenomenon is observed throughout the present investigation. However, the reduction of phenols at N5 and N10 is controversial in this study which may be due to the insufficient supplement of nanosilica to soil. Biocontrol agent is also essential for elevated level of phenols in plants.24 It is an interesting observation of increasing phenolic content and improved leaf physical strength to resist fungal stress in the present study. The crucial colonization level to be reached is estimated at 105–106 CFU g−1 of root in case of P. fluorescens treatment that protects plants from G. tritici or Pythium sp.13 The defense mechanisms in maize after the treatment of nanosilica and biocontrol agent are presented schematically as follows:
 | (1) |
 | (2) |
| SiO2 + P. fluorescens → better biocontrol + leaf erectness | (3) |
Our previous investigations on the role of nanosilica on conferring antagonistic action in maize against fungal pathogens reveal the changes in the response of stress responsive compounds to pathogenic infection.25 Similarly, present study is evident in developing plant protection by the combination of P. fluorescens and silica nanoparticles which further help to formulate novel biocontrol agents. A potent formulation can be developed for sustainable cropping from this biocomposite (SNPs and P. fluorescens). Hence, nanoscale silica acts as one of the mediators for defense reactions in planta compared to other conventional silica sources.
This investigation is aimed at providing a better biocontrol action against fungal pathogens as well as insects affecting maize crop. Nanosilica treated maize possesses rough leaf surface and good physical strength than that treated with bulk silica due to higher silica accumulation. A combination of P. fluorescens and SNPs in soil enhances phenolic activity and hence reduces the stress by the suppression of responsive enzymes in maize. This elevated level of phenols is found to induce silica accumulation in leaf epidermis, thereby conferring a protective physical barrier as well as induced disease resistance. Thus, one can formulate an effective biofertilizer/biocomposite for sustainable crop cultivation by using nanosilica.
We acknowledge the financial support provided by Defence Research and Development Organisation, New Delhi, India (ERIP/ER/0905113/M/01/1216) to carry out this research project.
Notes and references
- N. Debnath, S. Das, D. Seth, R. Chandra, S. C. Bhattacharya and A. Goswami, J. Pestic. Sci., 2011, 84, 99–105 Search PubMed.
- R. Suriyaprabha, G. Karunakaran, R. Yuvakkumar, V. Rajendran, P. Prabu and N. Kannan, J. Nanopart. Res., 2012, 14, 1–14 CrossRef.
- R. Yuvakkumar, V. Elango, V. Rajendran, N. Kannan and P. Prabu, Int. J. Green Nanotechnol., 2011, 3, 180–190 CrossRef CAS.
- L. R. Pokhrel and B. Dubey, Sci. Total Environ., 2013, 452–453, 321–332 CrossRef CAS PubMed.
- O. Yamamoto, Int. J. Inorg. Mater., 2001, 3, 643–646 CrossRef CAS.
- J. Peng, Y. Sun, Q. Liu, Y. Yang, J. Zhou, W. Feng, X. Zhang and F. Li, Nano Res., 2012, 5, 770–782 CrossRef CAS.
- V. Ghormade, M. V. Deshpande and K. M. Paknikar, Biotechnol. Adv., 2011, 29, 792–803 CrossRef CAS PubMed.
- S. Baruah and J. Dutta, Environ. Chem. Lett., 2009, 7, 161–204 CrossRef PubMed.
- E. Epstein, Silicon in Agriculture, Elsevier Science, Amsterdam, 2001, pp. 1–15 Search PubMed.
- J. F. Ma and N. Yamaji, Cell. Mol. Life Sci., 2008, 65, 3049–3057 CrossRef CAS PubMed.
- D. An, Y. Guo, Y. Zhu and Z. Wang, Chem. Eng. J., 2010, 162, 509–514 CrossRef CAS PubMed.
- T. K. Barik, B. Sahu and V. Swain, Parasitol. Res., 2008, 103, 253–258 CrossRef CAS PubMed.
- D. Haas and G. Defago, Nat. Rev. Microbiol., 2005, 3, 307–319 CrossRef CAS PubMed.
- A. Gamliel and J. Katan, Phytopathology, 1993, 83, 68–75 CrossRef.
- G. Hwang, I. S. Ahna, B. J. Mhin and J. Y. Kim, Colloids Surf., B, 2012, 97, 138–144 CrossRef CAS PubMed.
- B. B. Landa, D. M. Mavrodi, L. S. Thomashow and D. M. Weller, Phytopathology, 2003, 93, 982–994 CrossRef CAS PubMed.
- Z. Huang, X. Zheng, D. Yan, G. Yin, X. Liao, Y. Kang, Y. Yao, D. Huang and B. Hao, Langmuir, 2008, 24, 4140–4144 CrossRef CAS PubMed.
- C. Ulrichs, F. Krause, T. Rocksch, A. Goswami and I. Mewis, Commun. Agric. Appl. Biol. Sci., 2006, 71, 171–178 CAS.
- D. K. R. Robinson and G. Salejova-Zadrazilova, Observatory NANO, Working Paper Version, 2010.
- R. Suriyaprabha, G. Karunakaran, R. Yuvakkumar, V. Rajendran and N. Kannan, Curr. Nanosci., 2012, 8, 902–908 CrossRef CAS.
- G. Karunakaran, R. Suriyaprabha, P. Manivasakan, R. Yuvakkumar, V. Rajendran and N. Kannan, J. Nanosci. Nanotechnol., 2012, 13(1), 678–685 CrossRef PubMed.
- F. Fauteux, W. Remus-Borel, J. G. Menzies and R. R. Belanger, FEMS Microbiol. Lett., 2005, 249, 1–6 CrossRef CAS PubMed.
- E. Epstein, Ann. Appl. Biol., 2009, 155, 155–160 CrossRef CAS.
- D. Ghanmi, D. J. McNally, N. Benhamou, J. G. Menzies and R. R. Belanger, Physiol. Mol. Plant Pathol., 2004, 64, 189–199 CrossRef CAS PubMed.
- R. Suriyaprabha, G. Karunakaran, K. Kavitha, R. Yuvakkumar, V. Rajendran and N. Kannan, IET Nanobiotechnol., 2013, 1–5, DOI:10.1049/iet-nbt.2013.0004.
- V. Ramamoorthy, T. Raguchander and R. Samiyappan, Euro. J. Plant Pathol., 2002, 108, 429–441 CrossRef CAS.
- R. Hammerschmidt, E. M. Nuckles and C. J. Ku, Physiol. Plant Pathol., 1982, 20, 73–82 CrossRef CAS.
- K. Wakabayashi, T. Hoson and S. Kamisaka, Plant Physiol., 1997, 113, 967–973 CAS.
Footnotes |
| † Topographical (2D) and hardness measurements were carried out to explore the surface morphology and roughness of the maize leaves treated with SNPs and microparticles. The measurement was carried out using Ubi 1 Scanning Quasistatic nanoindentation (TI-700; Hysitron, MN, USA). A constant load of 1 μN was used to create an indentation with a Berkovich pyramidal indenter. Both the loading and unloading times were kept constant as 5 s at a rate of 200 nm s−1. |
| ‡ After 10 days of Pseudomonas sp. treatment, maize roots were collected and washed with sterile distilled water. The buffer extraction and estimation were carried out for plant defense enzymes such as phenylalanine ammonia lyase (PAL),26 peroxidase,27 and polyphenol oxidase (PPO)28 using spectrophotometer (U-2900; Hitachi, Japan). The enzyme activity was expressed in terms of changes in absorbance per minute. The total phenols from maize leaves of different treatments were extracted using methanol and estimated spectrophotometrically as the method described by Wakabayashi et al.28 |
| This journal is © The Royal Society of Chemistry 2014 |