 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceUnexpected similarities between the Schizosaccharomyces and human blood metabolomes, and novel human metabolites†
Romanas
Chaleckis
abc,
Masahiro
Ebe
c,
Tomáš
Pluskal
c,
Itsuo
Murakami
b,
Hiroshi
Kondoh
*b and
Mitsuhiro
Yanagida
*c
aGraduate School of Biostudies, Kyoto University, Sakyo-ku, Kyoto, Japan
bGraduate School of Medicine, Kyoto University, Sakyo-ku, Kyoto, Japan. E-mail: hkondoh@kuhp.kyoto-u.ac.jp
cOkinawa Institute of Science and Technology Graduate University (OIST), Onna-son, Okinawa, Japan. E-mail: myanagid@gmail.com
First published on 2nd July 2014
Abstract
Metabolomics, a modern branch of chemical biology, provides qualitative and quantitative information about the metabolic states of organisms or cells at the molecular level. Here we report non-targeted, metabolomic analyses of human blood, using liquid chromatography-mass spectrometry (LC-MS). We compared the blood metabolome to the previously reported metabolome of the fission yeast, Schizosaccharomyces pombe. The two metabolomic datasets were highly similar: 101 of 133 compounds identified in human blood (75%) were also present in S. pombe, and 45 of 57 compounds enriched in red blood cells (RBCs) (78%) were also present in yeast. The most abundant metabolites were ATP, glutathione, and glutamine. Apart from these three, the next most abundant metabolites were also involved in energy metabolism, anti-oxidation, and amino acid metabolism. We identified fourteen new blood compounds, eight of which were enriched in RBCs: citramalate, GDP-glucose, trimethyl-histidine, trimethyl-phenylalanine, trimethyl-tryptophan, trimethyl-tyrosine, UDP-acetyl-glucosamine, UDP-glucuronate, dimethyl-lysine, glutamate methyl ester, N-acetyl-(iso)leucine, N-acetyl-glutamate, N2-acetyl-lysine, and N6-acetyl-lysine. Ten of the newly identified blood metabolites were also detected in S. pombe, and ten of the 14 newly identified blood metabolites were methylated or acetylated amino acids. Trimethylated or acetylated free amino acids were also abundant in white blood cells. It may be possible to investigate their physiological roles using yeast genetics.
Introduction
Metabolomics is a rapidly growing branch of comprehensive, post-genomic, quantitative chemical biology.1–5 It aims to profile small molecules present in living organisms, and is now recognized as an important tool for studying metabolic regulation in a synthetic way, together with transcriptomic and proteomic analyses.6 Mass spectrometry (MS), a highly sensitive molecular detection method, can reveal thousands of intracellular metabolites over a very broad range of concentrations. However, the number of identified, named compounds is surprisingly small. One reason is the paucity of commercially available standards. Second, it is difficult to identify unknown ions, due to the limited structural information that can be obtained using MS. Third, the concentrations of many metabolites present in biological samples are very low, and they tend to undergo changes during sample preparation. Hence fewer than several hundred relatively abundant compounds are usually reported in comprehensive metabolomic studies.7–9Red blood cells (RBCs or erythrocytes) deliver oxygen to tissues and remove carbon dioxide. In humans, approximately 5 L of blood circulates through the whole body per minute.10 In healthy individuals, RBCs comprise 45% of the blood volume, and the remainder is mostly plasma, with a tiny additional fraction of leucocytes or white blood cells (WBCs). Plasma contains proteins (e.g. albumins, globulins, fibrinogen, etc.), electrolytes, hormones, glucose, and other metabolites. Mammalian RBCs exclude nuclei, mitochondria, and other prominent cell organelles, such as lysosomes, endoplasmic reticulum, and Golgi bodies during erythropoiesis in the bone marrow. Mature, simplified and specialized for gas transfer, RBCs have no demonstrable protein synthesis, nor tricarboxylic acid (TCA) cycle activity.11 Nevertheless, RBCs produce adenosine triphosphate (ATP) glycolytically, maintain redox homeostasis, and osmoregulate.12 Human RBCs have a relatively long life span of about 120 days.13 When senescent, they are captured by the spleen for degradation.
Numerous blood metabolomic analyses have been previously reported, mostly of serum and plasma.14–19 However, metabolomics of whole blood or RBCs have been less well investigated, except for several reports on long-term stored blood,20–23 blood of disease patients,24,25 or blood marker compounds of food intake.26 Reports on the WBC metabolome are scarce.27,28 This study focuses on the metabolome of blood cells, mainly RBCs, separated from plasma by low speed centrifugation. We also briefly investigated the metabolome of WBCs prepared by Ficoll gradient centrifugation, to compare it with that of RBCs.
The fission yeast, Schizosaccharomyces pombe, a eukaryotic microbe with a genome encoding only ∼5000 genes,29,30 is an excellent model organism to study nutritional control of the transition between proliferation and quiescence (e.g. Yanagida et al.31). We have employed metabolomic analysis of S. pombe cells in order to understand cellular metabolic states under different nutritional regimes and as governed by different genotypes. Initially, we reported the identification of 123 compounds using liquid chromatography-mass spectrometry (LC-MS) and their changes following heat stress and genetic perturbations.8 We described the accumulation of antioxidants, glutathione, and ergothioneine in a proteasome regulatory subunit mutant mts3-1.32 Also we made targeted measurements of intermediates of the coenzyme A biosynthetic pathway from pantothenate,33 investigated changes in the cellular metabolome upon glucose34 and nitrogen starvation,35 and accumulation of various metabolites in the quiescence-defective Δklf1 mutant.36 These studies showed that S. pombe may be used as a model organism to study comprehensive metabolic patterns under defined genetic and physiological conditions.
S. pombe cells grow and divide in culture media containing only 0.08% glucose as rapidly as in regular synthetic culture media containing 2% glucose; however, their division rate is impaired if extracellular glucose drops below 0.08%34 (4.4 mM), a concentration that is roughly equivalent to that in the blood of healthy humans before breakfast. That the glucose concentration required to sustain life is roughly equivalent between such diverse eukaryotes is remarkable. In this study we report further significant conservation in metabolites between evolutionarily distant human RBCs and S. pombe vegetative cells.
Results
Preparation and analysis of human blood metabolomic samples
Blood samples (4–5 mL) for metabolomic analysis were collected from healthy volunteers before breakfast (schematized in Fig. 1A, Materials and methods). WBCs stained with Giemsa (Fig. 1B) were infrequently observed (<1%) in blood. WBCs were not removed, since rapid quenching of metabolic reactions was necessary to obtain reproducible, quantitative data (Ficoll gradient centrifugation to separate WBCs is a time-consuming step). Low-molecular-weight metabolites were isolated by filtration with a 10 kDa cut-off filter at 4 °C, concentrated on a rotary evaporator, re-suspended in 40 μL 50% acetonitrile, and stored at −80 °C until analysis. | ||
| Fig. 1 Preparation and analysis of blood metabolomic samples. (A) Metabolic compounds were extracted in 50% MeOH at −40 °C from whole blood, plasma, and RBCs (Materials and methods). Extracted metabolites were isolated using a 10 kDa cut-off filter, concentrated by a rotary evaporator, and analyzed on an LC-MS system, as illustrated. (B) Blood cells stained with Giemsa solution under a microscope. Arrows indicate WBC. (C) 3D plots of raw LC-MS data of the plasma fraction (top) and the RBC fraction (bottom) obtained in positive ionization mode are shown: X-axis, retention time (RT, min); Y-axis, m/z; Z-axis, signal intensity. Twenty identified peaks are shown as examples. Peaks 1–10 are detected in both plasma and RBCs. These are amino acids, creatine, carnitine, dietary metabolites (caffeine, dimethyl-xanthine), and compounds introduced during sample preparation (HEPES as an internal standard, NH4Cl formed in the LC-MS system). Peaks 11–20 are enriched in RBC samples. Many compounds are involved in energy production, anti-oxidation, and amino acid metabolism (see text). | ||
For LC-MS analysis, metabolites were first separated by hydrophilic interaction chromatography (ZIC-pHILIC column; Merck SeQuant),37 and detected using an LTQ Orbitrap MS (Thermo Fisher Scientific) in full scan mode (100–1000 m/z, ratio of mass-to-charge) with both positive and negative electrospray ionization. LC-MS data contain semi-quantitative information about thousands of compounds in human blood. For compound analysis and quantification, we employed basically the same procedures used in previous analyses of S. pombe metabolites8 (Fig. 1C). For quantification, we integrated peak curves, obtaining peak areas in arbitrary AU units. ATP and glutathione are RBC-enriched, meaning that peak areas in the RBC fraction were at least 2-fold higher than the corresponding peaks in plasma (see ‘Fifty-seven RBC-enriched compounds’ below). It is difficult to obtain reproducible quantitative data on reduced glutathione (GSH) due to its auto-oxidation during sample preparation. For that reason, only levels of oxidized glutathione (GSSG) are reported in the present study.
Thirty-two compounds identified in human blood were not detected in yeast
MZmine 2 software38 was used for data processing and identification of blood metabolites. We employed an in-house database of m/z and RT values of compounds previously identified in fission yeast studies (ref. 8 and other references). For peaks not in the database, we performed a search using online databases HMDB,39 KEGG,40 or ChemSpider.41 Whenever possible, the identified compounds were verified using purchased standards. In some cases, isomers (e.g. N-acetyl-leucine, N-acetyl-isoleucine; paraxanthine, theobromine, theophylline) could not be clearly distinguished by LC and were designated by more general names (e.g., N-acetyl-(iso)leucine or dimethyl-xanthine, respectively). To identify metabolites for which standards were not available, we performed MS/MS analysis. Methyl-lysine, trimethyl-phenylalanine, and trimethyl-tyrosine were tentatively identified and described (Fig. S1A–C, ESI†). We were able to identify 133 compounds in blood, representing 14 categories (Table 1).| Category/compound | Status | In fission yeast | New blood component | RBC enrichedb | Ratio RBC![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :plasma ± standard deviationc :plasma ± standard deviationc |
Peak area,d blood | Peak area,d fission yeast |
|---|---|---|---|---|---|---|---|
| a One hundred thirty-three identified metabolic compounds detected in human blood metabolome samples by LC-MS. The status of the compounds was confirmed by either commercially available standards (STD) or MS/MS analysis (MS/MS). Compounds not detected in fission yeast (32 compounds) and compounds not reported as blood metabolites (14 compounds) are marked by ○ in respective columns. b On average in multiple persons. c Ratios between RBC and plasma samples calculated from four blood samples donated by the same person within 24 hours. Values and standard deviations rounded to 2 significant numbers. ND* – not detected. Values >2 shown in bold. d Peak areas defined as H, high (>108 AU); M, medium (107–108 AU); L, low (<107 AU); ND – not detected. In the case of blood, an equivalent of ∼5 μl of blood was injected into the LC-MS system, for fission yeast ∼0.75 μl of internal cell volume (see Materials and methods). | |||||||
| Nucleotides | |||||||
| ADP | STD | ○ | R | 64.0 ± 15.0 | M | M | |
| AMP | STD | ○ | R | 52.0 ± 6.8 | M | M | |
| ATP | STD | ○ | R | 81.0 ± 13.0 | H | H | |
| CDP | STD | ○ | R | 4.6 ± 2.1 | L | L | |
| CTP | STD | ○ | R | 2.2 ± 0.48 | L | M | |
| GDP | STD | ○ | R | 32.0 ± 5.5 | L | M | |
| GMP | STD | ○ | R | 3.5 ± 0.29 | L | L | |
| GTP | STD | ○ | R | 84.0 ± 14.0 | M | M | |
| IMP | STD | ○ | R | 180.0 ± 48.0 | L | L | |
| UDP | STD | ○ | R | 2.3 ± 0.5 | L | M | |
| UMP | STD | ○ | R | 19.0 ± 3.6 | L | L | |
| UTP | STD | ○ | R | 4.1 ± 1.6 | L | H | |
| Nucleosides, nucleobases and derivatives | |||||||
| 1-Methyl-adenosine | STD | ○ | 1.3 ± 0.14 | M | M | ||
| 1-Methyl-guanosine | STD | ○ | 1.1 ± 0.36 | L | L | ||
| Adenine | STD | ○ | 3.4 ± 0.055 | L | M | ||
| Adenosine | STD | ○ | 0.045± 0.012 | L | L | ||
| Caffeine | STD | 0.77 ± 0.057 | M | ND | |||
| Cytidine | STD | ○ | 2.1 ± 0.22 | L | L | ||
| Dimethyl-guanosine | STD | ○ | 0.77 ± 0.1 | L | L | ||
| Dimethyl-xanthine | STD | 0.88 ± 0.044 | M | ND | |||
| Guanosine | STD | ○ | 0.12 ± 0.077 | L | L | ||
| Hypoxanthine | STD | ○ | 1.4 ± 0.22 | L | L | ||
| Inosine | STD | ○ | 0.059 ± 0.12 | L | L | ||
| Uracil | STD | ○ | 1.0 ± 0.041 | L | L | ||
| Urate | STD | 0.69 ± 0.22 | M | ND | |||
| Uridine | STD | ○ | 1.0 ± 0.027 | L | L | ||
| Vitamins, coenzymes | |||||||
| 4-Aminobenzoate | STD | 2.5 ± 0.86 | L | ND | |||
| NAD+ | STD | ○ | R | 130.0 ± 30.0 | M | H | |
| NADH | STD | ○ | R | 80.0 ± 68.0 | L | M | |
| NADP+ | STD | ○ | R | 7200.0 ± 3000.0 | L | L | |
| NADPH | STD | ○ | R | 340.0 ± 220.0 | L | L | |
| Nicotinamide | STD | ○ | R | 26.0 ± 5.7 | M | M | |
| Pantothenate | STD | ○ | 3.3 ± 1.1 | L | M | ||
| Nucleotide-sugar derivatives | |||||||
| GDP-glucose | STD | ○ | ○ | R | 20.0 ± 3.6 | L | M |
| UDP-acetyl-glucosamine | STD | ○ | ○ | R | 11.0 ± 1.4 | L | M |
| UDP-glucose | STD | ○ | R | 35.0 ± 6.6 | M | H | |
| UDP-glucuronate | STD | ○ | R | 37.0 ± 6.2 | L | ND | |
| Sugar phosphates | |||||||
| 6-Phosphogluconate | STD | ○ | R | 150.0 ± 25.0 | L | L | |
| Diphospho-glycerate | STD | ○ | R | 1700.0 ± 350.0 | H | L | |
| Fructose-1,6-diphosphate | STD | ○ | R | 1400.0 ± 360.0 | M | M | |
| Fructose-6-phosphate | STD | ○ | R | 23.0 ± 2.2 | L | M | |
| Glucose-6-phosphate | STD | ○ | R | 32.0 ± 4.1 | M | M | |
| Glyceraldehyde-3-phosphate | STD | ○ | R | 910.0 ± 1100.0 | L | L | |
| Glycerol-2-phosphate | STD | ○ | R | 3.7 ± 1.1 | L | M | |
| Pentose-phosphate | STD | ○ | R | 39.0 ± 12.0 | L | M | |
| Phosphoenolpyruvate | STD | ○ | R | 1000.0 ± 780.0 | L | M | |
| Phosphoglycerate | STD | ○ | R | 150.0 ± 17.0 | M | M | |
| Sedoheptulose-7-phosphate | STD | ○ | R | 3.6 ± 0.76 | L | M | |
| Sugars and derivatives | |||||||
| 1,5-Anhydroglucitol | STD | 0.95 ± 0.14 | M | ND | |||
| Gluconate | STD | ○ | R | 16.0 ± 1.8 | M | L | |
| Glucosamine | STD | ○ | 0.89 ± 0.13 | M | L | ||
| Glucose | STD | ○ | 0.87 ± 0.12 | M | L | ||
| myo-Inositol | STD | ○ | 1.3 ± 0.39 | L | L | ||
| N-Acetyl-D-glucosamine | STD | ○ | R | 35.0 ± 1.4 | M | L | |
| Quinic acid | STD | ○ | 2.1 ± 2.0 | L | M | ||
| Organic acids | |||||||
| 2-Oxoglutarate | STD | ○ | 0.94 ± 0.16 | L | L | ||
| Chenodeoxycholate | STD | 0.49 ± 0.14 | M | ND | |||
| cis-Aconitate | STD | ○ | 0.33 ± 0.11 | L | L | ||
| Citramalate | STD | ○ | ○ | R | 3.2 ± 0.87 | L | M |
| Citrate | STD | ○ | 0.28 ± 0.069 | M | M | ||
| Fumarate | STD | ○ | R | 5.6 ± 0.9 | L | L | |
| Glutarate | STD | ○ | 0.94 ± 0.072 | L | L | ||
| Glycerate | STD | 0.69 ± 0.18 | L | ND | |||
| Malate | STD | ○ | R | 4.6 ± 0.6 | L | L | |
| Succinate | STD | ○ | 0.92 ± 0.21 | L | L | ||
| Standard amino acids | |||||||
| Arginine | STD | ○ | 0.57 ± 0.072 | H | H | ||
| Asparagine | STD | ○ | 1.6 ± 0.18 | L | L | ||
| Aspartate | STD | ○ | R | 6.3 ± 0.91 | L | M | |
| Glutamate | STD | ○ | R | 3.4 ± 0.74 | M | H | |
| Glutamine | STD | ○ | 0.56 ± 0.053 | H | H | ||
| Histidine | STD | ○ | 0.93 ± 0.054 | M | H | ||
| Isoleucine | STD | ○ | 0.95 ± 0.14 | M | L | ||
| Leucine | STD | ○ | 1.1 ± 0.098 | M | L | ||
| Lysine | STD | ○ | 0.91 ± 0.15 | L | L | ||
| Methionine | STD | ○ | 0.76 ± 0.099 | M | L | ||
| Phenylalanine | STD | ○ | 0.93 ± 0.16 | H | M | ||
| Proline | STD | ○ | 0.92 ± 0.054 | H | L | ||
| Serine | STD | ○ | 1.1 ± 0.19 | L | L | ||
| Threonine | STD | ○ | 1.1 ± 0.12 | M | M | ||
| Tryptophan | STD | ○ | 0.47 ± 0.054 | M | M | ||
| Tyrosine | STD | ○ | 1.1 ± 0.083 | M | M | ||
| Valine | STD | ○ | 0.87 ± 0.07 | M | L | ||
| Methylated amino acids | |||||||
| Betaine | STD | ○ | 1.2 ± 0.11 | H | L | ||
| Butyro-betaine | STD | R | 5.3 ± 0.52 | M | ND | ||
| Dimethyl-arginine | STD | ○ | 0.95 ± 0.12 | L | L | ||
| Dimethyl-lysine | STD | ○ | ○ | 0.51 ± 0.09 | L | L | |
| Dimethyl-proline (stachydrine) | STD | R | 4.9 ± 0.66 | H | ND | ||
| Methyl-histidine | STD | ○ | 0.92 ± 0.056 | L | M | ||
| Methyl-lysine | MS/MS | ○ | 0.67 ± 0.11 | M | L | ||
| S-Methyl-ergothioneine | STD | R | 1500.0 ± 2300.0 | L | ND | ||
| Trimethyl-histidine (hercynine) | MS/MS | ○ | ○ | R | 110.0 ± 140.0 | L | M |
| Trimethyl-lysine | STD | ○ | R | 6.3 ± 1.5 | M | L | |
| Trimethyl-phenylalanine | MS/MS | ○ | R | 14.0 ± 9.0 | L | ND | |
| Trimethyl-tryptophan (hypaphorine) | STD | ○ | R | 6.1 ± 1.7 | H | ND | |
| Trimethyl-tyrosine | MS/MS | ○ | R | ND* | L | ND | |
| Acetylated amino acids | |||||||
| N-Acetyl-(iso)leucine | STD | ○ | ○ | 1.8 ± 0.39 | L | L | |
| N-Acetyl-arginine | STD | ○ | 0.34 ± 0.047 | L | M | ||
| N-Acetyl-aspartate | STD | ○ | 0.84 ± 0.2 | L | L | ||
| N-Acetyl-glutamate | STD | ○ | ○ | 0.82 ± 0.18 | L | M | |
| N-Acetyl-ornithine | STD | ○ | 2.6 ± 1.2 | L | L | ||
| N 2-Acetyl-lysine | STD | ○ | ○ | 2.1 ± 0.3 | L | L | |
| N 6-Acetyl-lysine | STD | ○ | ○ | 0.88 ± 0.26 | L | M | |
| Other amino acid derivatives | |||||||
| 2-Aminoadipate | STD | ○ | 0.91 ± 0.073 | L | L | ||
| Arginino-succinate | STD | ○ | 0.69 ± 0.46 | L | M | ||
| Citrulline | STD | ○ | 0.97 ± 0.19 | M | M | ||
| Creatine | STD | R | 7.0 ± 1.6 | H | ND | ||
| Creatinine | STD | 0.97 ± 0.079 | H | ND | |||
| Glutamate methyl ester | STD | ○ | ○ | 1.5 ± 0.28 | L | L | |
| Hippurate | STD | 0.54 ± 0.083 | M | ND | |||
| Histamine | STD | ○ | 1.0 ± 0.5 | L | L | ||
| Indoxyl-sulfate | STD | 0.43 ± 0.092 | M | ND | |||
| Kynurenine | STD | 0.41 ± 0.081 | L | ND | |||
| Ornithine | STD | ○ | 1.2 ± 0.16 | L | M | ||
| Phosphocreatine | STD | R | 3.0 ± 0.95 | L | ND | ||
| Quinolinic acid | STD | 0.35 ± 0.04 | L | ND | |||
| S-Adenosyl-homocysteine | STD | ○ | R | 2100.0 ± 1600.0 | L | M | |
| S-Adenosyl-methionine | STD | ○ | R | 57.0 ± 62.0 | L | L | |
| Taurine | STD | 0.92 ± 0.18 | M | ND | |||
| Carnitines | |||||||
| Acetyl-carnitine | STD | R | 4.0 ± 0.58 | H | ND | ||
| Butyryl-carnitine | STD | 0.95 ± 0.2 | M | ND | |||
| Carnitine | STD | 0.85 ± 0.1 | H | ND | |||
| Decanoyl-carnitine | STD | 0.42 ± 0.11 | M | ND | |||
| Dodecanoyl-carnitine | STD | 0.51 ± 0.072 | L | ND | |||
| Hexanoyl-carnitine | STD | 0.68 ± 0.087 | L | ND | |||
| Isovaleryl-carnitine | STD | 0.94 ± 0.25 | L | ND | |||
| Octanoyl-carnitine | STD | 0.45 ± 0.086 | M | ND | |||
| Propionyl-carnitine | STD | R | 5.3 ± 0.82 | M | ND | ||
| Tetradecanoyl-carnitine | STD | R | 11.0 ± 15.0 | L | ND | ||
| Choline derivatives | |||||||
| CDP-choline | STD | ○ | 0.98 ± 0.37 | L | L | ||
| Glycerophosphocholine | STD | ○ | 1.5 ± 0.24 | M | H | ||
| Antioxidant | |||||||
| Ergothioneine | STD | ○ | R | 100.0 ± 4.8 | H | M | |
| Glutathione disulfide (GSSG) | STD | ○ | R | 1900.0 ± 430.0 | H | H | |
| Ophthalmic acid | STD | ○ | R | 310.0 ± 150.0 | L | M | |
To compare compounds present in S. pombe with those in human blood, S. pombe cells were cultivated at 26 °C in synthetic, minimal EMM2 medium with 0.1% glucose (5.6 mM; roughly the same glucose concentration found in blood). Among 133 identified blood metabolites, 32 were not found in fission yeast (Table 1). They include three nucleoside bases and derivatives, one coenzyme precursor (4-aminobenzoate), one nucleotide-sugar derivative (UDP-glucuronate), one sugar derivative (1,5-anhydroglucitol), two organic acids (chenodeoxycholic acid, glyceric acid), six methylated amino acids including dimethyl-proline, eight other amino acid derivatives including creatine, creatinine, and taurine, and ten carnitines. Thus, 24 of 32 compounds are derived from three categories, namely, methylated amino acids, other amino acids, and carnitines. These three categories illustrate the major difference between human blood and S. pombe metabolomes. Some compounds may play specific roles in higher eukaryotes. For example, creatine and phosphocreatine are stored in muscles as an energy source, and creatinine is the metabolic byproduct of creatine phosphate.42 Carnitines are produced in the liver and stored in muscles for consumption and transport of fatty acids.43 Urate is a purine metabolic byproduct, high blood concentrations of which can cause gout,44 while caffeine is a xanthine alkaloid and of dietary origin.45,46S. pombe does not produce caffeine, but can become hypersensitive or resistant to caffeine by certain mutations.47,48
Fourteen newly identified blood compounds
To our knowledge, 14 metabolites have not hitherto been reported in human blood, based on a recent report of detected blood metabolites49 and literature database searches (Table 1). These new blood metabolites include citramalate, dimethyl-lysine, GDP-glucose, glutamate methyl ester, N-acetyl-glutamate, N-acetyl-(iso)leucine, N2-acetyl-lysine, N6-acetyl-lysine, trimethyl-histidine, trimethyl-phenylalanine, trimethyl-tryptophan, trimethyl-tyrosine, UDP-acetyl-glucosamine and UDP-glucuronate. The eight compounds in boldface were enriched in RBCs, while the ten compounds in italics were also found in S. pombe. Ten of the 14 novel blood metabolites are methylated or acetylated amino acids.Quantification of blood metabolite peaks
Each blood sample produced thousands of peaks in positive and negative ionization modes with a broad range (104–109 AU) of peak areas. We quantified compounds on the basis of their peak areas: high (H, over 108 AU), medium (M, 107–108 AU) and low (L, <107 AU). In blood samples, L, M, and H groups comprised 92, 7, and 1% of all peaks, respectively.Quantitative reproducibility of peak areas was examined by collecting two blood samples independently from the same person at 1 h intervals. Each pair of samples (#1 and #2) of blood, plasma, and RBCs was compared in a scatter plot (Fig. 2A and Fig. S2A, ESI†). In all cases, 85–87% of peak areas varied less than 2-fold (0.5–2.0×). Fission yeast samples obtained under identical conditions showed similar reproducibility.8 Very small peaks (area <106 AU) showed larger deviations. For 133 compounds identified in blood, plasma or RBCs, however, 97% of peaks in the compared samples changed less than 2-fold (Fig. 2B and Fig. S2B, ESI†). Thus, in both blood and fission yeast metabolomes, quantitative reproducibility was better for identified peaks.8
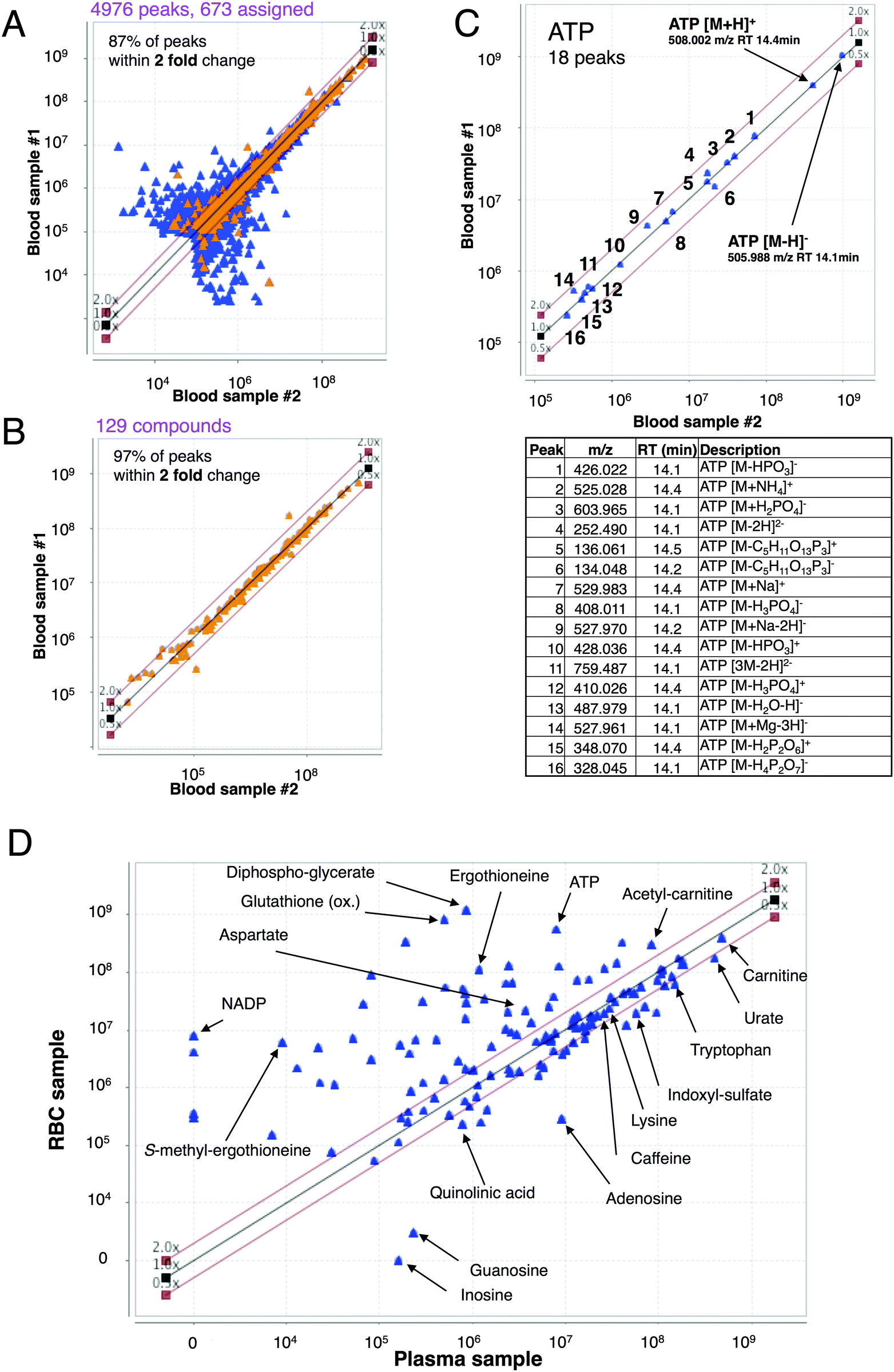 | ||
| Fig. 2 Quantification of peak area reproducibility examined by a scatter plot. Blood was donated twice by the same person in 1 h, and the two blood samples were processed separately (samples #1 and #2). (A) Scatter plot of all peaks detected in both blood samples (positive and negative ionization modes combined). 87% of peaks differed less than 2-fold. Less than 15% of these peaks could be assigned to a known compound (assigned peaks marked yellow). (B) A scatter plot of 129 identified compounds detected in blood samples #1 and #2. Approximately 97% of these peaks were found within 2-fold change. (C) Multiple peaks are produced by abundant compounds such as ATP. In addition to the primary singly charged ions (indicated by arrows), compounds produce multiply charged ions, fragments of the molecule, adducts with salts, and complexes with other compounds eluting at similar retention times. We were able to identify 18 peaks related to ATP, listed in the inset table. (D) RBC and plasma samples were prepared from the same person, and the peak areas were determined for identified compounds, and compared in a scatter plot. | ||
Highly abundant metabolites form various adducts or fragments, resulting in multiple MS peaks. For quantification, we used singly charged proton adducts in positive [M + H]+ and negative modes [M − H]−. ATP produced these two peaks as its highest signals (Fig. 2C). ATP also produced 16 additional peaks (6 in positive and 10 in negative mode). Since their retention time (RT) was basically identical to that of the corresponding primary peak, we suspect that these additional peaks were produced during ionization in the MS. For ergothioneine, 17 peaks were identified in addition to the primary peaks (Fig. S2C, ESI†).
In blood samples we were able to identify 37 (74%) peaks in group H, 118 (33%) peaks in group M, and 518 peaks (11%) in group L. The total number of assigned peaks (673) is much larger than that of actually identified compounds (133), due to the fact that many metabolites produced multiple peaks. A number of peaks were also produced by electrolytes such as NH4Cl, originating from NaCl in blood samples. While several thousand peaks were obtained by LC-MS, the actual number of compounds that can be detected in blood may be much less, possibly ∼1000.
Fifty-seven RBC-enriched compounds
To determine the degree of compound enrichment in RBCs vs. plasma, samples of both were prepared from the same blood donor several times. We designate RBC-enriched compounds as those having an RBC:plasma ratio more than 2.0 (Fig. 2D and Table 1; detailed annotation of all peaks in Fig. S3, ESI†).
ATP and glutathione showed particularly large peak areas (>108 AU) in the RBC sample, but much smaller in the plasma sample (RBC:plasma ratios of 81 and 1900, respectively). In contrast, carnitine and urate showed RBC:plasma ratios of 0.85 and 0.69, respectively, even though their peak areas were large (>108 AU) in both samples.
Fifty-seven compounds were enriched in RBCs (Table 1). Most metabolites highly enriched in RBC fractions (RBC:plasma ratio >30) were nucleotides (ADP, AMP, ATP, GDP, GTP, IMP), sugar phosphates (6-phosphogluconate, diphospho-glycerate, fructose-1,6-diphosphate, glyceraldehyde-3-phosphate, pentose-phosphate, phosphoenolpyruvate, phosphoglycerate), vitamins (NAD+, NADH, NADP+, NADPH), antioxidants (ergothioneine, glutathione disulfide (GSSG), ophthalmic acid), methylated compounds (S-adenosyl-homocysteine, S-methyl-ergothioneine, tetradecanoyl-carnitine, trimethyl-histidine) and N-acetyl-D-glucosamine. There was no evidence of leakage of these highly enriched compounds from RBCs, confirming that cells were not damaged during handling. Metabolites moderately enriched in RBCs (RBC:plasma ratio between 2 and 30) contain compounds from the categories mentioned above, as well as nucleotide-sugar derivatives and at least one or two compounds from other categories (except antioxidants and choline derivatives).
Many RBC-enriched compounds, such as nucleotides and sugar-phosphates, are involved in RBC metabolic pathways.23 Others such as acetyl-carnitine50 and trimethyl-lysine51 were previously reported to be enriched in RBCs, but their origins and roles in RBCs are unknown. Interestingly, eight of the fourteen newly discovered blood components (nucleotide-sugar derivatives and methylated amino acids) were also enriched in RBCs.
In contrast to RBC-enriched compounds, adenosine, guanosine, and inosine were scarcely detected in RBCs. Thus, brief centrifugation to sediment RBCs in combination with LC-MS quantification enabled us to classify metabolites into several groups based upon their RBC:plasma ratios (>30 highly RBC-enriched, <30, >2 RBC-enriched; <2 present in both plasma and RBC).
Comparison of semi-quantified metabolites between blood and S. pombe
Some compounds exhibited similar abundance in blood and S. pombe cells (Table 1). ATP (H), ADP (M), AMP (M) and GTP (M) were similar in both, whereas UTP was abundant in S. pombe (H), but not in blood (L). Except for UTP, these nucleotides are enriched in RBC; thus RBCs evidently have a low requirement for UTP, since RNA synthesis does not occur in RBCs. Diphospho-glycerate, an allosteric regulator of hemoglobin present at mM concentrations in RBCs,52 was highly abundant (H) in blood, but not in S. pombe (L). Pentose-phosphate and sedoheptulose-7-phosphate were more abundant in S. pombe (M) than in blood (L), indicating that growing and dividing yeast cells require greater amounts of pentose phosphate pathway intermediates.Seventeen regular and seven acetylated amino acids were commonly present in blood and yeast. Fourteen methylated amino acids were significantly different between blood and S. pombe, while seven methylated amino acids (betaine, dimethyl-arginine, dimethyl-lysine, methyl-histidine, methyl-lysine, trimethyl-histidine, trimethyl-lysine) were commonly present in both, and seven others (butyro-betaine, dimethyl-proline, S-methyl-ergothioneine, trimethyl-phenylalanine, trimethyl-tryptophan, trimethyl-tyrosine) were detected only in human samples. Three anti-oxidants, glutathione, ergothioneine, and ophthalmic acid, were present in both human and yeast samples.
Eighteen S. pombe compounds not detected in human blood
Eighteen compounds present in S. pombe were not detected in RBCs or in plasma (Table S1, ESI†). Compounds that control fast cell division and growth, such as cyclic AMP (an activator of protein kinase A53), AICAR (5′-phosphoribosyl-5-amino-4-imidazolecarboxamide; an activator of AMP-dependent protein kinase, AMPK54), SAICAR (succinylaminoimidazolecarboxamide ribose-5′-phosphate; an activator of pyruvate kinase, PKM255), and PRPP (phosphoribosyl pyrophosphate, involved in nucleotide metabolism56), are present in S. pombe, but not in blood. Acetyl-CoA, biotin, coenzyme A, FAD (flavin adenine dinucleotide), and HMG-CoA may be required for rapid cell division. Ferrichrome is a cyclic hexapeptide present in lower eukaryotes, such as Schizosaccharomyces and Ustilago, which forms a complex with iron.57 Trehalose is a disaccharide having an α,α-1,1-glucosidic bond implicated in anhydrobiotic (anti-desiccant) and anti-oxidant mechanisms.58 Trehalose is not synthesized in the human body. Trehalose-6-phosphate has been implicated in plant flowering,59 but no function in fission yeast has been proposed. Saccharopine is an intermediate of lysine metabolism.60Compounds detected in WBCs in comparison with those from RBCs and S. pombe
We then examined metabolites of WBCs isolated by Ficoll gradient centrifugation (Table S2, ESI†). While the metabolic profile of WBCs was similar to that of RBCs, seven metabolites, 3′-5′-cAMP, acetyl-CoA, CMP, CoA, FAD (flavin adenine dinucleotide), HMG-CoA, and PRPP, were detected only in WBCs (Table S1, ESI†). Differences between the two are mainly the types of vitamins and coenzymes. 4-Aminobenzoate was detected in blood and RBCs, but not in WBCs. Acetyl-CoA, biotin, FAD, and HMG-CoA, described above, belong to the coenzyme category. Seven compounds, which seem to be mainly involved in cell growth and division in eukaryotes, were found in both WBCs and S. pombe (Table S1, ESI†).Summary and categorization of detected blood compounds
Human blood metabolites identified in this study are summarized in Fig. 3. Three nucleosides, adenosine, guanosine and inosine, may be restricted to plasma, as RBC:plasma ratios are close to zero (Table 1). cis-Aconitate (0.3), citrate (0.3), indoxyl-sulfate (0.4), kynurenine (0.4), N-acetyl-arginine (0.3), and quinolinic acid (0.4) appear to be found primarily in plasma. In contrast, NADP+, S-adenosyl-homocysteine, phosphoenolpyruvate, glutathione disulfide (GSSG), diphospho-glycerate, and fructose-1,6-diphosphate were highly enriched in RBC fractions. Their RBC:plasma ratios are rather large (>1000). RBC compounds of intermediate abundance (ratio >50–<1000) included 6-phosphogluconate, ADP, AMP, ATP, ergothioneine, GTP, IMP, NAD+, NADPH, ophthalmic acid, phosphoglycerate, S-methyl-ergothioneine, tetradecanoyl-carnitine, and trimethyl-histidine. All other compounds are presumably present in both RBCs and plasma.
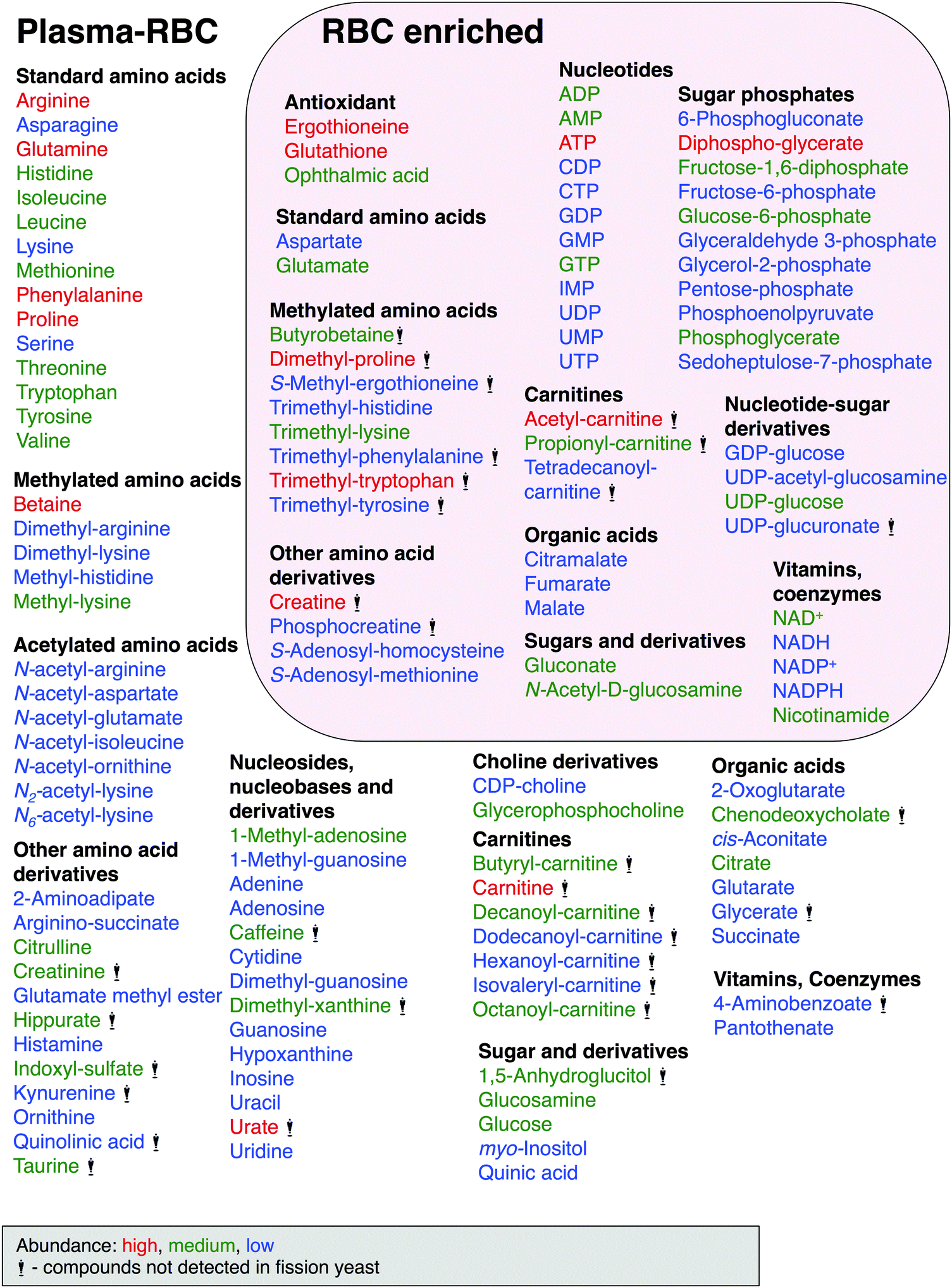 | ||
| Fig. 3 Human blood compounds identified and characterized in the present study. Compounds that were either RBC-enriched (57) or not (76), based on whether the ratios of their RBC:plasma peak areas were either >2 or <2, respectively (Table 1). Abundance of compounds classified by peak area size, indicated by color, red (high), green (medium) and blue (low). Compounds with the statue symbol are not present in S. pombe. See text for details. | ||
Energy and anti-oxidant metabolites abundant in human blood and fission yeast
Some compounds (ATP, glutathione and glutamine) were highly abundant in both RBCs and S. pombe (Fig. 4). Most compounds in the high and medium categories are implicated in energy metabolism, anti-oxidation, or amino acid metabolism. In the WBC metabolome (Table S2, ESI†), ATP, glutathione, and glycerophosphocholine (instead of glutamine) were abundant. Some medium-level compounds (e.g. NAD+ and UDP-glucose) are required for production of high-energy compounds such as ATP, GTP, and UTP. Ergothioneine and ophthalmic acid are thought to be anti-oxidants. It thus appears that energy metabolites, anti-oxidants, and amino acid metabolites may be the most highly conserved in eukaryotes. | ||
| Fig. 4 Relatively high-abundance compounds in human blood and fission yeast. Compound abundance in human blood and S. pombe. For example, “high–high” indicates that ATP, glutathione, and glutamine are highly abundant in both blood and S. pombe. See text for details. | ||
Discussion
In the present study we performed LC-MS-based metabolomic analysis of human blood, plasma, and RBCs in comparison with previously published metabolomic results from the fission yeast, S. pombe.8,34 Analysis of the S. pombe metabolome was performed simultaneously, and results were consistent with those of previous reports. This comparative study enabled us to learn which metabolomic features are conserved between these distantly related cellular systems. The LC column we employed can separate hydrophilic compounds with high resolution, but is not appropriate for separating hydrophobic compounds; therefore, our data contain limited information on lipids, except for two choline derivatives, CDP-choline and glycerophosphocholine. For this reason, conclusions regarding metabolite conservation should be restricted to hydrophilic compounds.Thirty-two compounds were found in blood, but not in S. pombe (Fig. 3). Metabolite compositions of blood and fission yeast are unexpectedly similar, with 75% identified compound in blood also present in fission yeast. The WBC metabolome is also highly similar to those of RBCs and yeast. However, metabolites mostly belonging to three categories (ten carnitines, six methylated amino acids, eight other amino acid derivatives) were not detected in S. pombe (Table 1). If we compare metabolites in the remaining 11 categories, only 8 of 89 compounds were not present in S. pombe (caffeine, dimethyl-xanthine, urate, 4-aminobenzoate, UDP-glucuronate, 1,5-anhydroglucitol, chenodeoxycholic acid, and glyceric acid); therefore the overall similarity between the two metabolomes is 90% (81/89 × 100). Differences are mostly restricted to carnitines and amino acid derivatives.
Among the fifteen high abundance blood compounds, however, S. pombe lacks six (40%) metabolites (acetyl-carnitine, carnitine, creatine, dimethyl-proline, trimethyl-tryptophan and urate), indicating that the most abundant compounds in blood are quite different from those of S. pombe. Conversely, among the 14 high abundance S. pombe compounds, only 2 (trehalose, ferrichrome) were not present in human blood, indicating that 86% are also found in human blood. We presume that the high abundance metabolites of S. pombe may be more ‘fundamental’ than those of blood, since S. pombe is a single-celled eukaryote. Blood contains a multitude of specialized metabolites. Carnitine is not strictly a metazoan compound, as it is also synthesized from lysine in Neurospora.61 However, a great variety of carnitine derivatives (9) carrying different fatty acids might be a feature of higher eukaryotes.
The high similarity of these two metabolomes raises the possibility that S. pombe genetics might be useful to understand the role of certain metabolites, such as small anti-oxidants (ophthalmic acid, ergothioneine, and glutathione), which are enzymatically synthesized in S. pombe. S-Adenosyl-homocysteine (SAH) and S-adenosyl-methionine (SAM), coenzymes involved in the methionine cycle, were also enriched in RBCs. S-Adenosyl-methionine synthase has been reported in RBCs.62 In RBCs, SAM has been shown to act as a coenzyme for protein carboxyl methylation,63 synthesis of phosphatidylcholine,64 and N-, O- and S-methyltransferase activities (reviewed by Weinshilboum et al.65). It remains to be determined how SAH and SAM in RBCs are involved in the production of methylated compounds. We are particularly interested in determining whether the levels of free methylated amino acids are controlled by SAM and SAH. To this end, we isolated a number of S. pombe mutants of methionine cycle enzymes and their metabolic profiles are being examined (T. Hayashi et al., unpublished results).
We found 14 novel human blood compounds, eight of which were methylated or N-acetylated amino acids. To our knowledge, there has been no report describing these as blood components. Blood data presented in this report came from four healthy volunteers. We further extended our analysis to more than 30 people (Chaleckis et al., manuscript in preparation), and these compounds were universally present, suggesting that their occurrence is neither accidental nor specific to certain dietary customs. Interestingly, eight of these compounds (UDP-acetyl-glucosamine, citramalate, dimethyl-lysine, trimethyl-histidine, N-acetyl-glutamate, N2-acetyl-lysine, N6-aceltyl-glutamate and glutamate methyl ester) were also present in S. pombe. Their physiological roles can thus be further investigated using S. pombe as a model. Seven of the eight compounds are RBC-enriched. RBCs may require UDP-acetyl-glucosamine, a nucleotide sugar and a coenzyme, as a signaling molecule of sugar metabolism, like in other eukaryotic cells, including S. pombe.66 UDP-glucuronic acid is synthesized in the liver, binds to hormones or toxic compounds, and is also used for the synthesis of glucuronic acid-containing polysaccharides.67,68 It is not present in lower eukaryotes. Citramalate is an intermediate in bacterial glutamate degradation.69 Citramalate and glutamate methyl-ester are also present in S. pombe, but their physiological role is not understood. Two acetylated amino acids, N2-acetyl-lysine and N6-acetyl-lysine, are present in plasma and RBCs, as well as in S. pombe, but their biological role is little understood. N-acetyl-glutamate is involved in the removal of waste from the body in the urine as it is an allosteric cofactor of carbamoyl phosphate synthetase I, the first enzyme in the urea cycle.70
Five novel blood compounds are methylated amino acids, four (histidine, phenylalanine, tryptophan, tyrosine) of which are trimethylated and enriched in RBCs. Trimethyl-histidine (hercynine) is a precursor of ergothioneine, but humans do not synthesize ergothioneine; therefore this compound might be of dietary origin or possibly a degradation product of ergothioneine,71 whereas S. pombe can produce it.34,35 Trimethyl-tryptophan (hypaphorine) was highly abundant in RBCs. It has been reported that this compound has soporific72 and anti-glycemic effects in mice.73 Hypaphorine was reported in human milk following maternal consumption of legumes.74
ATP and glutathione were selectively enriched in RBCs (scarcely present in plasma), while glutamine was found in both plasma and RBCs in roughly equal amounts. Eleven sugar phosphate compounds required for sugar and energy metabolism were all found in RBC-enriched fractions and also in S. pombe. Similarly, all twelve nucleotides, four nucleotide-sugar derivatives, and five coenzyme NAD-related compounds were selectively enriched in RBCs. Fifty-six percent of RBC-enriched compounds are energy-related; these compounds are also found in S. pombe. Three anti-oxidant compounds, glutathione, ergothioneine, and ophthalmic acid, were enriched in RBCs and abundant in S. pombe. Glutathione and ophthalmic acid may be synthesized in RBCs, as the synthetic enzymes encoded by the human genes are present in RBCs.75 Aspartate and glutamate were selectively enriched in RBCs. Both are excitatory neurotransmitters. Inhibitory transmitters, GABA and glycine, are difficult to measure using our method. Glutamate may be partly utilized for the synthesis of glutathione.76 These energy and anti-oxidant compounds are most likely essential for maintaining RBCs during their relatively long lifespan of 120 days, and these compounds are also common to S. pombe.
Materials and methods
Chemicals and reagents
Standards for metabolite confirmation were obtained from various vendors (Table S3, ESI†), depending on the compound. Ergothioneine and S-methyl-ergothioneine were a kind gift from Tetrahedron, France.Human subject characteristics
Three healthy male and one female volunteers (23–33 years old) participated in this study. Blood samples for metabolomic analysis were taken in the morning and subjects were asked not to eat breakfast to ensure at least 12 h of fasting prior to sampling.Ethics statement
Written informed consent was obtained from all donors in accordance with the Declaration of Helsinki. All experiments were performed in compliance with relevant Japanese laws and institutional guidelines. All protocols were approved by the Ethical Committee on Human Research of Kyoto University Hospital and by the Human Subjects Research Review Committee of the Okinawa Institute of Science and Technology Graduate University (OIST).Blood sample preparation for metabolomic analysis
Blood samples for metabolomic analysis were drawn by venipuncture into 5 mL heparinized tubes (Terumo). Immediately, 0.2 mL blood samples (8–12 × 108 RBC) were quenched in 1.8 mL of 55% methanol at −40 °C. The remainder of each blood sample was centrifuged at 120 × g for 15 min at room temperature to separate plasma and RBCs. After centrifugation, 0.2 mL of separated plasma and RBCs (14–20 × 108 RBC), respectively, was quenched in 1.8 mL of 55% methanol at −40 °C. Ten nmol each of HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) and PIPES (piperazine-N,N′-bis(2-ethanesulfonic acid)) was added to each sample to serve as standards. After brief vortexing, the samples were transferred to Amicon Ultra 10 kDa cut-off filters (Millipore, Billerica, MA, USA) to remove proteins and cell debris. Following sample concentration by vacuum evaporation, each sample was re-suspended in 40 μL of 50% acetonitrile and 1 μL (equivalent to 5 μL of initial sample) was used for each injection into the LC-MS system.Isolation of leucocytes by Ficoll gradient centrifugation
For isolation of leucocytes we employed Ficoll gradient centrifugation. Blood of two donors, 20 mL each, was collected into heparinized tubes (Terumo) and subsequently diluted with the same volume of PBS buffer. Diluted blood was carefully layered onto the same volume of Lympholyte-H (Cosmo Bio, Tokyo, Japan) and centrifuged at room temperature for 20 min at 800 × g. Separated leucocytes were washed 3× in PBS (1 min, 350 × g, 4 °C). After dilution, 0.2 mL of leucocytes (∼5 × 107 cells) was quenched in 1.8 mL of 55% methanol at −40 °C. After the addition of HEPES and PIPES, samples were processed as described above. Metabolites were isolated from 10 μL of leukocyte cell volume (assuming an individual leukocyte cell volume of 200 fL77 and an estimated 5 × 107 cells per sample). One μL of the metabolome sample was injected into the LC-MS system, corresponding to ∼0.25 μL of leukocyte intracellular volume.Fission yeast growth conditions and preparation for metabolomics analysis
The wild-type heterothallic haploid 972 h−S. pombe strain78 was used for metabolomic experiments. Cells were cultivated at 26 °C in minimal synthetic medium EMM279 with 0.1% glucose (5.6 mM) content. Limited glucose media were prepared by mixing regular EMM2 (2% glucose) medium with EMM2-G (0% glucose) in an appropriate ratio. Preparation of fission yeast samples was done as described previously.8 Metabolites were isolated from 30 μL of total S. pombe cell volume (S. pombe cell diameter 3.5 μm;80 cells grown at 5.6 mM glucose have lengths of 13 μm,34 and thus using a formula from Mitchison,80 the mean cell volume is 120 fL; total number of cells per sample is 2.5 × 108). LC-MS sample injections (1 μL) corresponded to ∼0.75 μL of S. pombe intracellular volume.LC-MS analysis
LC-MS data were obtained using a Paradigm MS4 HPLC system (Michrom Bioresources, Auburn, CA, USA) coupled to an LTQ Orbitrap mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA) as described previously.8 Briefly, LC separation was performed on a ZIC-pHILIC column (Merck SeQuant, Umeå, Sweden; 150 mm × 2.1 mm, 5 μm particle size). Acetonitrile (A) and 10 mM ammonium carbonate buffer (pH 9.3; B) were used as the mobile phase, with gradient elution from 80% A to 20% A in 30 min, at a flow rate of 100 μL mL−1. Peak areas for metabolites of interest were integrated using MZmine 2 software version 2.10.38 Detailed data analytical procedures and parameters have been described previously.8 Metabolites were initially identified by searching their m/z values in on-line databases (see text) or by predicting chemical formulae from mass spectra.81 Identified peaks were verified by analyzing pure standards (STD category) to confirm their retention times, or in cases where pure standards were not available, by analyzing their fragmentation patterns (MS/MS category).Abbreviations
| RBC | Red blood cell |
Funding
This work was supported by grants from the Okinawa Intellectual Cluster Program (to MY, HK), from the Ministry of Education, Culture, Sports, Science, and Technology of Japan, and from Japan Science and Technology Agency (to HK). We acknowledge the generous support from the Okinawa Institute of Science and Technology Graduate University. RC was supported by Japanese government pre-doctoral scholarship (Monbukagakusho). IM was supported by a postdoctoral fellowship from the Okinawa Intellectual Cluster Program. Funding agencies had no role in study design, data collection and analysis, the decision to publish, or preparation of the manuscript.Disclosure statement
The authors declare that they have no competing interests.Acknowledgements
We thank Dr Masayuki Kobayashi for help with WBC isolation and Eri Shibata for providing excellent technical assistance. We are most grateful to Dr Steve Aird for editing and helpful discussion during preparation of the present paper.References
- A. R. Fernie, R. N. Trethewey, A. J. Krotzky and L. Willmitzer, Nat. Rev. Mol. Cell Biol., 2004, 5, 763–769 CrossRef CAS PubMed.
- D. B. Kell, Curr. Opin. Microbiol., 2004, 7, 296–307 CrossRef CAS PubMed.
- R. Goodacre, S. Vaidyanathan, W. B. Dunn, G. G. Harrigan and D. B. Kell, Trends Biotechnol., 2004, 22, 245–252 CrossRef CAS PubMed.
- J. K. Nicholson and J. C. Lindon, Nature, 2008, 455, 1054–1056 CrossRef CAS PubMed.
- G. J. Patti, O. Yanes and G. Siuzdak, Nat. Rev. Mol. Cell Biol., 2012, 13, 263–269 CrossRef CAS PubMed.
- M. Y. Hirai, M. Yano, D. B. Goodenowe, S. Kanaya, T. Kimura, M. Awazuhara, M. Arita, T. Fujiwara and K. Saito, Proc. Natl. Acad. Sci. U. S. A., 2004, 101, 10205–10210 CrossRef CAS PubMed.
- M. J. Brauer, J. Yuan, B. D. Bennett, W. Lu, E. Kimball, D. Botstein and J. D. Rabinowitz, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 19302–19307 CrossRef CAS PubMed.
- T. Pluskal, T. Nakamura, A. Villar-Briones and M. Yanagida, Mol. Biosyst., 2010, 6, 182–198 RSC.
- T. Soga, Y. Ohashi, Y. Ueno, H. Naraoka, M. Tomita and T. Nishioka, J. Proteome Res., 2003, 2, 488–494 CrossRef CAS.
- A. C. Allison, Nature, 1960, 188, 37–40 CrossRef CAS.
- S. M. Rapoport, T. Schewe and B.-J. Thiele, Blood Cell Biochemistry, 1990, vol. 1, pp. 151–194 Search PubMed.
- R. van Wijk and W. W. van Solinge, Blood, 2005, 106, 4034–4042 CrossRef CAS PubMed.
- R. S. Franco, Am. J. Hematol., 2009, 84, 109–114 CrossRef PubMed.
- A. Jiye, J. Trygg, J. Gullberg, A. I. Johansson, P. Jonsson, H. Antti, S. L. Marklund and T. Moritz, Anal. Chem., 2005, 77, 8086–8094 CrossRef PubMed.
- K. A. Lawton, A. Berger, M. Mitchell, K. E. Milgram, A. M. Evans, L. Guo, R. W. Hanson, S. C. Kalhan, J. A. Ryals and M. V. Milburn, Pharmacogenomics, 2008, 9, 383–397 CrossRef CAS PubMed.
- S. J. Bruce, I. Tavazzi, V. Parisod, S. Rezzi, S. Kochhar and P. A. Guy, Anal. Chem., 2009, 81, 3285–3296 CrossRef CAS PubMed.
- T. Kimura, Y. Noguchi, N. Shikata and M. Takahashi, Curr. Opin. Clin. Nutr. Metab. Care, 2009, 12, 49–53 CrossRef CAS PubMed.
- N. J. Serkova, T. J. Standiford and K. A. Stringer, Am. J. Respir. Crit. Care Med., 2011, 184, 647–655 CrossRef CAS PubMed.
- N. Psychogios, D. D. Hau, J. Peng, A. C. Guo, R. Mandal, S. Bouatra, I. Sinelnikov, R. Krishnamurthy, R. Eisner, B. Gautam, N. Young, J. Xia, C. Knox, E. Dong, P. Huang, Z. Hollander, T. L. Pedersen, S. R. Smith, F. Bamforth, R. Greiner, B. McManus, J. W. Newman, T. Goodfriend and D. S. Wishart, PLoS One, 2011, 6, e16957 CAS.
- A. D'Alessandro, G. M. D'Amici, S. Vaglio and L. Zolla, Haematologica, 2012, 97, 107–115 CrossRef PubMed.
- A. D'Alessandro, F. Gevi and L. Zolla, Mol. Biosyst., 2013, 9, 1196–1209 RSC.
- T. Nishino, A. Yachie-Kinoshita, A. Hirayama, T. Soga, M. Suematsu and M. Tomita, PLoS One, 2013, 8, e71060 CAS.
- T. Nishino, A. Yachie-Kinoshita, A. Hirayama, T. Soga, M. Suematsu and M. Tomita, J. Biotechnol., 2009, 144, 212–223 CrossRef CAS PubMed.
- D. Darghouth, B. Koehl, J. F. Heilier, G. Madalinski, P. Bovee, G. Bosman, J. Delaunay, C. Junot and P. H. Romeo, Haematologica, 2011, 96, 1861–1865 CrossRef CAS PubMed.
- D. Darghouth, B. Koehl, G. Madalinski, J. F. Heilier, P. Bovee, Y. Xu, M. F. Olivier, P. Bartolucci, M. Benkerrou, S. Pissard, Y. Colin, F. Galacteros, G. Bosman, C. Junot and P. H. Romeo, Blood, 2011, 117, e57–e66 CrossRef CAS PubMed.
- U. Catalan, M. A. Rodriguez, M. R. Ras, A. Macia, R. Mallol, M. Vinaixa, S. Fernandez-Castillejo, R. M. Valls, A. Pedret, J. L. Griffin, R. Salek, X. Correig, M. J. Motilva and R. Sola, Mol. Biosyst., 2013, 9, 1411–1422 RSC.
- D. Y. Sze and O. Jardetzky, Biochim. Biophys. Acta, 1990, 1054, 181–197 CrossRef CAS.
- R. Lee and P. Britz-McKibbin, Anal. Chem., 2009, 81, 7047–7056 CrossRef CAS PubMed.
- V. Wood, R. Gwilliam, M. A. Rajandream, M. Lyne, R. Lyne, A. Stewart, J. Sgouros, N. Peat, J. Hayles, S. Baker, D. Basham, S. Bowman, K. Brooks, D. Brown, S. Brown, T. Chillingworth, C. Churcher, M. Collins, R. Connor, A. Cronin, P. Davis, T. Feltwell, A. Fraser, S. Gentles, A. Goble, N. Hamlin, D. Harris, J. Hidalgo, G. Hodgson, S. Holroyd, T. Hornsby, S. Howarth, E. J. Huckle, S. Hunt, K. Jagels, K. James, L. Jones, M. Jones, S. Leather, S. McDonald, J. McLean, P. Mooney, S. Moule, K. Mungall, L. Murphy, D. Niblett, C. Odell, K. Oliver, S. O'Neil, D. Pearson, M. A. Quail, E. Rabbinowitsch, K. Rutherford, S. Rutter, D. Saunders, K. Seeger, S. Sharp, J. Skelton, M. Simmonds, R. Squares, S. Squares, K. Stevens, K. Taylor, R. G. Taylor, A. Tivey, S. Walsh, T. Warren, S. Whitehead, J. Woodward, G. Volckaert, R. Aert, J. Robben, B. Grymonprez, I. Weltjens, E. Vanstreels, M. Rieger, M. Schafer, S. Muller-Auer, C. Gabel, M. Fuchs, A. Dusterhoft, C. Fritzc, E. Holzer, D. Moestl, H. Hilbert, K. Borzym, I. Langer, A. Beck, H. Lehrach, R. Reinhardt, T. M. Pohl, P. Eger, W. Zimmermann, H. Wedler, R. Wambutt, B. Purnelle, A. Goffeau, E. Cadieu, S. Dreano, S. Gloux, V. Lelaure, S. Mottier, F. Galibert, S. J. Aves, Z. Xiang, C. Hunt, K. Moore, S. M. Hurst, M. Lucas, M. Rochet, C. Gaillardin, V. A. Tallada, A. Garzon, G. Thode, R. R. Daga, L. Cruzado, J. Jimenez, M. Sanchez, F. del Rey, J. Benito, A. Dominguez, J. L. Revuelta, S. Moreno, J. Armstrong, S. L. Forsburg, L. Cerutti, T. Lowe, W. R. McCombie, I. Paulsen, J. Potashkin, G. V. Shpakovski, D. Ussery, B. G. Barrell and P. Nurse, Nature, 2002, 415, 871–880 CrossRef CAS PubMed.
- V. Wood, M. A. Harris, M. D. McDowall, K. Rutherford, B. W. Vaughan, D. M. Staines, M. Aslett, A. Lock, J. Bahler, P. J. Kersey and S. G. Oliver, Nucleic Acids Res., 2012, 40, D695–D699 CrossRef CAS PubMed.
- M. Yanagida, N. Ikai, M. Shimanuki and K. Sajiki, Philos. Trans. R. Soc. London, Ser. B, 2011, 366, 3508–3520 CrossRef CAS PubMed.
- K. Takeda, T. Yoshida, S. Kikuchi, K. Nagao, A. Kokubu, T. Pluskal, A. Villar-Briones, T. Nakamura and M. Yanagida, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 3540–3545 CrossRef CAS PubMed.
- T. Nakamura, T. Pluskal, Y. Nakaseko and M. Yanagida, Open Biol., 2012, 2, 120117 CrossRef PubMed.
- T. Pluskal, T. Hayashi, S. Saitoh, A. Fujisawa and M. Yanagida, FEBS J., 2011, 278, 1299–1315 CrossRef CAS PubMed.
- K. Sajiki, T. Pluskal, M. Shimanuki and M. Yanagida, Metabolites, 2013, 3, 1118–1129 CrossRef CAS PubMed.
- M. Shimanuki, L. Uehara, T. Pluskal, T. Yoshida, A. Kokubu, Y. Kawasaki and M. Yanagida, PLoS One, 2013, 8, e78545 CAS.
- Y. Guo and S. Gaiki, J. Chromatogr. A, 2005, 1074, 71–80 CrossRef CAS PubMed.
- T. Pluskal, S. Castillo, A. Villar-Briones and M. Oresic, BMC Bioinf., 2010, 11, 395 CrossRef PubMed.
- D. S. Wishart, T. Jewison, A. C. Guo, M. Wilson, C. Knox, Y. Liu, Y. Djoumbou, R. Mandal, F. Aziat, E. Dong, S. Bouatra, I. Sinelnikov, D. Arndt, J. Xia, P. Liu, F. Yallou, T. Bjorndahl, R. Perez-Pineiro, R. Eisner, F. Allen, V. Neveu, R. Greiner and A. Scalbert, Nucleic Acids Res., 2013, 41, D801–D807 CrossRef CAS PubMed.
- M. Kanehisa and S. Goto, Nucleic Acids Res., 2000, 28, 27–30 CrossRef CAS.
- H. E. Pence and A. Williams, J. Chem. Educ., 2010, 87, 1123–1124 CrossRef CAS.
- R. J. Snow and R. M. Murphy, Mol. Cell. Biochem., 2001, 224, 169–181 CrossRef CAS.
- A. M. Evans and G. Fornasini, Clin. Pharmacokinet., 2003, 42, 941–967 CrossRef CAS PubMed.
- B. N. Ames, R. Cathcart, E. Schwiers and P. Hochstein, Proc. Natl. Acad. Sci. U. S. A., 1981, 78, 6858–6862 CrossRef CAS.
- J. J. Barone and H. R. Roberts, Food Chem. Toxicol., 1996, 34, 119–129 CrossRef CAS.
- N. L. Benowitz, Annu. Rev. Med., 1990, 41, 277–288 CrossRef CAS PubMed.
- H. Ohkura, Y. Adachi, N. Kinoshita, O. Niwa, T. Toda and M. Yanagida, EMBO J., 1988, 7, 1465–1473 CAS.
- K. Kumada, M. Yanagida and T. Toda, Mol. Gen. Genet., 1996, 250, 59–68 CAS.
- S. M. Rappaport, D. K. Barupal, D. Wishart, P. Vineis and A. Scalbert, Environ. Health Perspect., 2014 DOI:10.1289/ehp.1308015.
- M. B. Cooper, C. A. Forte and D. A. Jones, Biochim. Biophys. Acta, 1988, 959, 100–105 CrossRef CAS.
- M. Mizobuchi, M. Miyake, A. Sano and Y. Kakimoto, Biochim. Biophys. Acta, 1990, 1033, 119–123 CrossRef CAS.
- A. Joshi and B. O. Palsson, J. Theor. Biol., 1990, 142, 69–85 CrossRef CAS.
- Y. M. Yamashita, Y. Nakaseko, I. Samejima, K. Kumada, H. Yamada, D. Michaelson and M. Yanagida, Nature, 1996, 384, 276–279 CrossRef CAS PubMed.
- J. M. Corton, J. G. Gillespie, S. A. Hawley and D. G. Hardie, Eur. J. Biochem., 1995, 229, 558–565 CrossRef CAS PubMed.
- K. E. Keller, I. S. Tan and Y. S. Lee, Science, 2012, 338, 1069–1072 CrossRef CAS PubMed.
- A. W. Murray, Annu. Rev. Biochem., 1971, 40, 811–826 CrossRef CAS PubMed.
- M. Schrettl, G. Winkelmann and H. Haas, BioMetals, 2004, 17, 647–654 CrossRef CAS.
- A. D. Elbein, Y. T. Pan, I. Pastuszak and D. Carroll, Glycobiology, 2003, 13, 17R–27R CrossRef CAS PubMed.
- V. Wahl, J. Ponnu, A. Schlereth, S. Arrivault, T. Langenecker, A. Franke, R. Feil, J. E. Lunn, M. Stitt and M. Schmid, Science, 2013, 339, 704–707 CrossRef CAS PubMed.
- H. Xu, B. Andi, J. Qian, A. H. West and P. F. Cook, Cell Biochem. Biophys., 2006, 46, 43–64 CrossRef CAS.
- C. J. Rebouche and H. P. Broquist, J. Bacteriol., 1976, 126, 1207–1214 CAS.
- K. L. Oden and S. Clarke, Biochemistry, 1983, 22, 2978–2986 CrossRef CAS.
- A. F. Perna, D. Ingrosso, V. Zappia, P. Galletti, G. Capasso and N. G. De Santo, J. Clin. Invest., 1993, 91, 2497–2503 CrossRef CAS PubMed.
- F. Hirata and J. Axelrod, Proc. Natl. Acad. Sci. U. S. A., 1978, 75, 2348–2352 CrossRef CAS.
- R. M. Weinshilboum, D. M. Otterness and C. L. Szumlanski, Annu. Rev. Pharmacol. Toxicol., 1999, 39, 19–52 CrossRef CAS PubMed.
- K. E. Wellen and C. B. Thompson, Nat. Rev. Mol. Cell Biol., 2012, 13, 270–276 CAS.
- R. H. Tukey and C. P. Strassburg, Annu. Rev. Pharmacol. Toxicol., 2000, 40, 581–616 CrossRef CAS PubMed.
- B. Mulloy and M. J. Forster, Glycobiology, 2000, 10, 1147–1156 CrossRef CAS PubMed.
- H. A. Barker, Annu. Rev. Biochem., 1981, 50, 23–40 CrossRef CAS PubMed.
- L. Caldovic and M. Tuchman, Biochem. J., 2003, 372, 279–290 CrossRef CAS PubMed.
- I. K. Cheah and B. Halliwell, Biochim. Biophys. Acta, 2012, 1822, 784–793 CrossRef CAS PubMed.
- M. Ozawa, K. Honda, I. Nakai, A. Kishida and A. Ohsaki, Bioorg. Med. Chem. Lett., 2008, 18, 3992–3994 CrossRef CAS PubMed.
- K. Chand, Akanksha, N. Rahuja, D. P. Mishra, A. K. Srivastava and R. Maurya, Med. Chem. Res., 2011, 20, 1505–1508 CrossRef CAS.
- B. O. Keller, B. T. Wu, S. S. Li, V. Monga and S. M. Innis, J. Agric. Food Chem., 2013, 61, 7654–7660 CrossRef CAS PubMed.
- A. Hirono, H. Iyori, I. Sekine, J. Ueyama, H. Chiba, H. Kanno, H. Fujii and S. Miwa, Blood, 1996, 87, 2071–2074 CAS.
- S. Whillier, B. Garcia, B. E. Chapman, P. W. Kuchel and J. E. Raftos, FEBS J., 2011, 278, 3152–3163 CrossRef CAS PubMed.
- G. B. Segel, G. R. Cokelet and M. A. Lichtman, Blood, 1981, 57, 894–899 CAS.
- H. Gutz, H. Heslot, U. Leupold and N. Loprieno, in Handbook of Genetics, ed. R. King, 1974, pp. 395–446 Search PubMed.
- S. L. Forsburg and N. Rhind, Yeast, 2006, 23, 173–183 CrossRef CAS PubMed.
- J. M. Mitchison, Exp. Cell Res., 1957, 13, 244–262 CrossRef CAS.
- T. Pluskal, T. Uehara and M. Yanagida, Anal. Chem., 2012, 84, 4396–4403 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Raw LC-MS data in mzML format were submitted to the MetaboLights repository (URL: http://www.ebi.ac.uk/metabolights). The accession number for the fission yeast and human blood metabolome comparison is MTBLS87, while that for metabolomic samples of a single individual's blood donated 4 times within 24 h to determine RBC:plasma metabolite distribution is MTBLS88. See DOI: 10.1039/c4mb00346b |
| This journal is © The Royal Society of Chemistry 2014 |