 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Uptake of self-secreted flavins as bound cofactors for extracellular electron transfer in Geobacter species†
Akihiro
Okamoto
a,
Koichiro
Saito
a,
Kengo
Inoue
b,
Kenneth H.
Nealson
c,
Kazuhito
Hashimoto
*a and
Ryuhei
Nakamura
*d
aDepartment of Applied Chemistry, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-8656, Japan. E-mail: hashimoto@light.t.u-tokyo.ac.jp
bInterdisciplinary Research Organization, University of Miyazaki, 5200 Kihara, Kiyotake, Miyazaki 889-1692, Japan
cDepartments of Earth Sciences and Biological Sciences, University of Southern California, Los Angeles, CA 90089, USA
dBiofunctional Catalysts Research Team, RIKEN Center for Sustainable Resource Science, 2-1 Hirosawa, Wako, Saitama 351-0198, Japan. E-mail: ryuhei.nakamura @riken.ac.jp
First published on 12th February 2014
Abstract
Geobacter species are among the most efficient current-producing bacterial species, yet their electron-transfer mechanisms have been scarcely investigated at the molecular level. Here, we provide evidence that Geobacter cells secrete and utilize riboflavin as a bound-cofactor in outer-membrane c-type cytochromes. This finding highlights the potential roles of riboflavin as a major electron carrier in current production.
Broader contextGeobacter sulfurreducens is an iron-reducing bacterium that has a significant content of c-type cytochromes (c-Cyts) in an outer-cell membrane (OM) and bacterial filaments, drawing keen attention as a model microorganism for the research of microbial fuel cells and bioremediation technology. Concerned with the ability of Geobacter to transport electrons to electrodes in microbial fuel cells, processes termed extracellular electron transport (EET), a great deal of research has been focused on identifying the molecular mechanisms behind the bacterial current generation. Herein, we report the first evidence that Geobacter utilizes self-secreted flavins as a redox cofactor in OM c-Cyts. Using a highly sensitive voltammetry technique, the key redox signal for the Geobacter EET was identified in vivo. Experiments using a mutant unable to produce OM c-Cyts, together with the spectroscopic and LC-MS analyses, revealed that the noble redox signal is assigned to the bound riboflavin associated with OM c-Cyts containing reduced hemes. As the heme redox state reflects a balance between the electron input from respiration and the output by EET, the present study not only significantly contributes to the molecular understanding of EET, but also highlights the microbial capability of utilizing self-secreted riboflavin as a regulator for intracellular redox homeostasis. |
Electron transfer from cell metabolic systems to exterior solid substrates, termed extracellular electron transfer (EET), is an intriguing aspect of microbial respiration.1,2 In anaerobic environments, particularly biofilms, EET is a terminal step of catabolism and is involved in redox sensing3 and intercellular4/interspecies5 energy transfer. It is also a fundamental process in microbial communities involved in energy production (e.g., microbial fuel cells),6 bioremediation of waste waters and contaminated sediments,7 and anaerobic pipeline corrosion.8
Over the last decade, several mechanisms for microbial EET have been proposed, including indirect electron transfer via redox-active organic electron shuttles9,10 and direct electron transfer by c-type cytochromes (c-Cyts) located in the outer membrane (OM)11 or on nanometer-scale bacterial filaments.12,13 In indirect EET, microbial cells are able to perform electron transfer without the necessity of direct contact with solid-phase electron acceptors. Thereby, as the surface area of the solids is limited, EET via self-secreted, naturally occurring or artificially supplemented soluble electron shuttles appears to be a major pathway for electron transfer to solids.10 However, such soluble redox molecules are apparently not involved in the EET process of Geobacter sulfurreducens,14 which is the most efficient current-producing microorganism characterized to date. This conclusion was reached after it was shown that the exchange of supernatants to fresh medium in an electrochemical reactor did not impair current production by G. sulfurreducens as it did in other microorganisms.14 Recently, we reported that cell-secreted redox molecules, such as flavins, have a high affinity for OM c-Cyts with reduced hemes in Shewanella oneidensis MR-1.15 This flavin binding to OM c-Cyt scaffolds was found to facilitate a one-electron redox reaction via semiquinone, resulting in a 103- to 105-fold enhancement of the EET rate compared to free flavin.15 Based on this finding, we speculated that Geobacter cells might also use self-secreted redox molecules as redox cofactors to promote EET. If true, the excreted molecules should not act as a diffusion-based electron shuttle in G. sulfurreducens, and thereby the supernatant exchange could not impair current production by G. sulfurreducens.14 This possibility is supported by the fact that, similar to S. oneidensis MR-1,16G. sulfurreducens have genes for both flavin biosynthesis and secretion (Fig. S1†), although their functions have not been confirmed.
In this paper, we report that G. sulfurreducens secrete flavin that contributes to EET at the cell/electrode interface, highlighting the crucial role of flavin redox cycling for efficient EET in this species.
To directly examine the secretion of flavin by G. sulfurreducens, a supernatant solution of anaerobically grown cultures was subjected to spectroscopic measurements (Fig. 1). Geobacter cells were grown in a defined medium supplemented with acetate (20 mM) as an electron donor and fumarate (80 mM) as an electron acceptor. As shown in Fig. 1a and b, both the peak intensities of emission at 525 nm and excitation at 370 and 440 nm increased when the growth curve of cells was in a sigmoidal phase. The emission and excitation spectral peaks were identical to those observed in the spectra of riboflavin (RF) and flavin mononucleotide (FMN) solutions without cells (Fig. S2a and b†), suggesting that G. sulfurreducens cells secrete flavin species during growth. In support of these data, mass chromatography patterns of the cell culture supernatant measured in selective ion monitoring (SIM) mode at m/z 375.35 identified that RF was present at a concentration of 100 nM, although FMN at m/z 455.34 was under the detection limit (Fig. 1c and S3†). These findings provide evidence that G. sulfurreducens secrete RF at concentrations comparable to that observed in cultures of anaerobically grown S. oneidensis MR-1.17
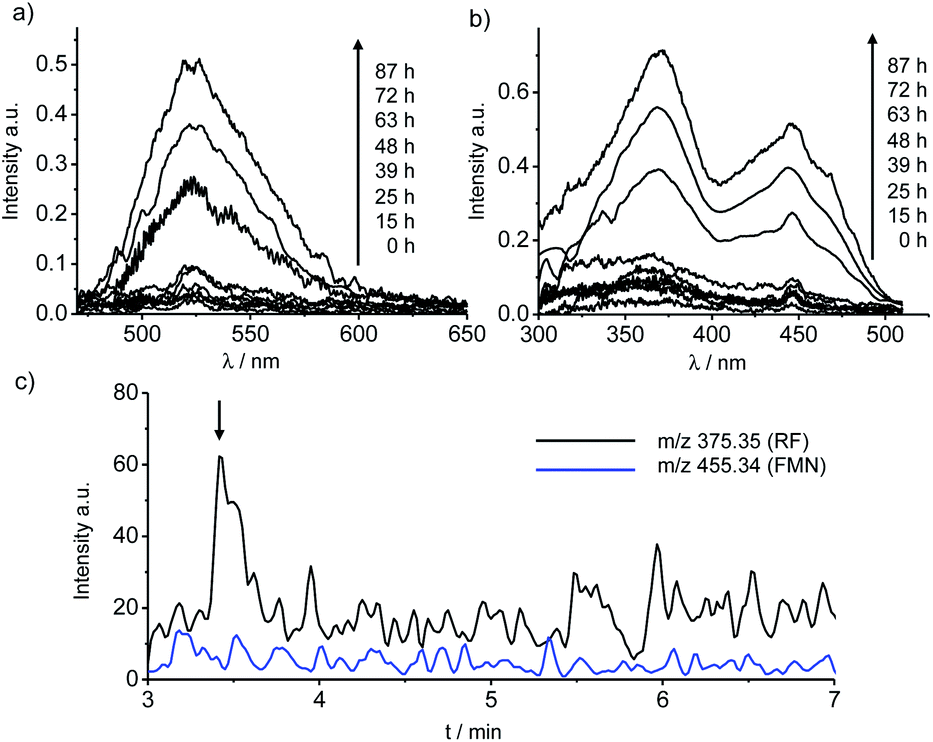 | ||
| Fig. 1 Detection of flavins secreted by G. sulfurreducens during growth in anaerobic medium. Emission (a) and excitation (b) spectra of the cell culture supernatant after 0, 15, 25, 39, 48, 63, 72, 87 h of incubation. (c) Mass chromatography patterns of the cell culture supernatant after 87 h of anaerobic culture. RF and FMN were detected by selected ion monitoring mode at 375.35 and 455.34 m/z, respectively. | ||
The contribution of secreted RF for EET was examined by adding a concentrated RF solution during the electrochemical cultivation of G. sulfurreducens. Current production (Ic) was measured using a cell suspension with an optical density at λ = 600 nm (OD600) of 0.2 cultured with 10 mM acetate as the sole electron source on an indium-tin oxide (ITO) electrode at a poised potential of +0.4 V (vs. SHE) (ESI†). At several time points during the course of microbial current production, differential pulse (DP) voltammetry was conducted (Fig. 2a). Three peaks were observed in DP voltammograms after 46 h (Fig. 2b), but only the redox peak current at −0.2 V (I−0.2) increased with time, whereas the peak current of the other redox peaks decreased inversely with time (Fig. 2b). When 1.0 μM RF was added to Geobacter cells on the electrode during current production under the same conditions as above, the Ic rapidly increased by ∼10% (Fig. S4†). Furthermore, as the addition of RF also caused an increase in I−0.2 (Fig. S5†), it is possible to assign the redox peak at −0.2 V to the redox cycling of RF. This result also indicates that Geobacter cells utilize self-secreted RF for EET, even before the addition of exogenous RF.
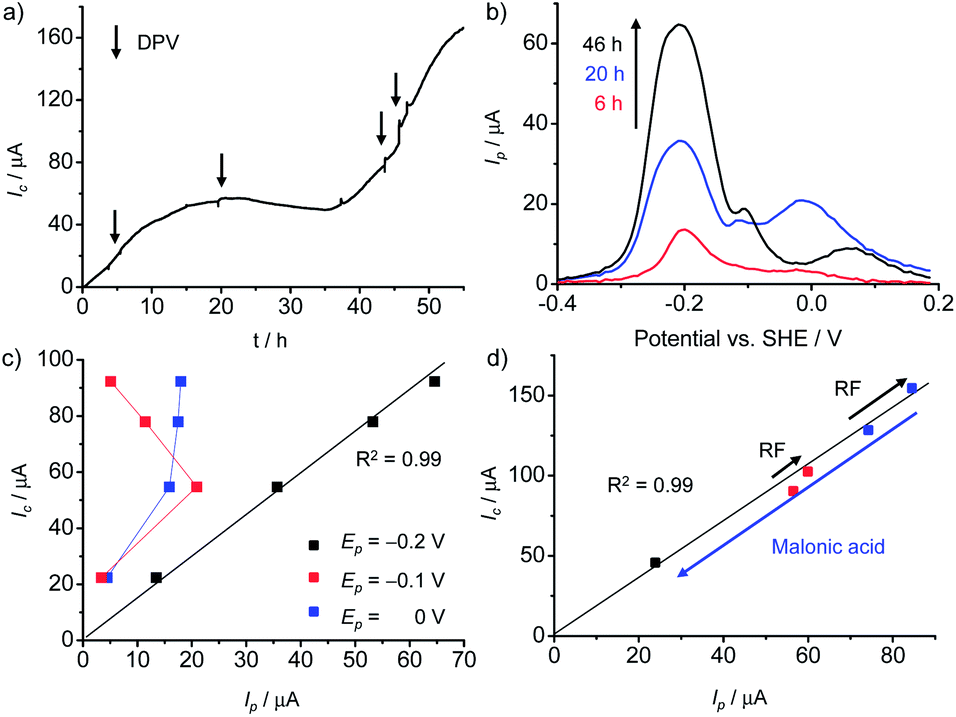 | ||
| Fig. 2 Role of riboflavin (RF) in extracellular electron transfer (EET) by G. sulfurreducens. (a) Current (Ic) versus time (t) measurements for microbial current generation by G. sulfurreducens on an ITO electrode. Arrows indicate the time when differential pulse (DP) voltammetry was measured. (b) DP voltammograms measured at t = 6, 20, and 46 h in panel a. (c) Plot of Ic in panel a against the peak current (Ip) at peak potentials (Ep) of −0.2, −0.1 and 0 V. (d) Plot of Ic against Ip at an Ep of −0.2 V before and after the addition of 1 μM RF and 100 mM malonic acid. The squares of the correlation coefficients were estimated by the addition of the point of origin to the obtained data. | ||
We plotted I−0.2 against Ic to quantify the contribution of the redox species at −0.2 V to EET. As shown in Fig. 2, Ic exhibited a positive correlation with I−0.2, as a fitted line passed through the point of origin with a high correlation coefficient (r2 = 0.998). Importantly, this trend was observed both before and after the addition of RF, and even after the addition of malonic acid as a metabolic inhibitor (Fig. 2d and S4†). These results confirm the assignment of the peak potential (Ep) at −0.2 V to the redox cycling of RF, and demonstrate that respiratory current generation by G. sulfurreducens is strongly coupled with the amount of electrochemically active RF. In addition, FMN exhibited a similar effect on the Ic and peak current to that seen with RF, but at a different Ep (Fig. S6†). Upon the addition of FMN, both the peak current at −0.175 V (I−0.175) and Ic increased, and a fitted line for the plot of I−0.175 and Ic passed through the point of origin (Fig. S6†), demonstrating the capability of G. sulfurreducens to utilize FMN as an efficient electron carrier. Together, these data provide evidence for the involvement of RF and/or FMN as important electron carriers at the interface between Geobacter cells and ITO electrodes.
The measured Ep in the DP voltammogram of RF and FMN solutions in the absence of cells significantly differ from those observed in Geobacter biofilms (Fig. S7†), suggesting that both flavins alter their redox properties, as is reported for flavins bound to OM c-Cyts in S. oneidensis MR-1.15 Assuming one of the c-Cyts located on bacterial filaments or embedded in the OM interacts with flavins as bound cofactors in G. sulfurreducens, insight into the location of c-Cyts that bind RF is important to determine the predominant EET pathway in this species. Fig. 3 shows scanning electron microscopy (SEM) images of G. sulfurreducens cells attached to the ITO electrode after 50 h of electrode cultivation. Neither filament-like assemblages nor multilayer biofilms were formed on the electrode surface, corresponding to a previous report where thick biofilms of G. sulfurreducens cells with Cyt-bound filaments required more than 4 days of electrochemical cultivation.18 Therefore, it appears that the EET process mediated by the redox cycling of flavins observed in Fig. 2 is the consequence of the activation of c-Cyts located primarily on the OM surface, as opposed to those located on the surface of conductive bacterial filaments.
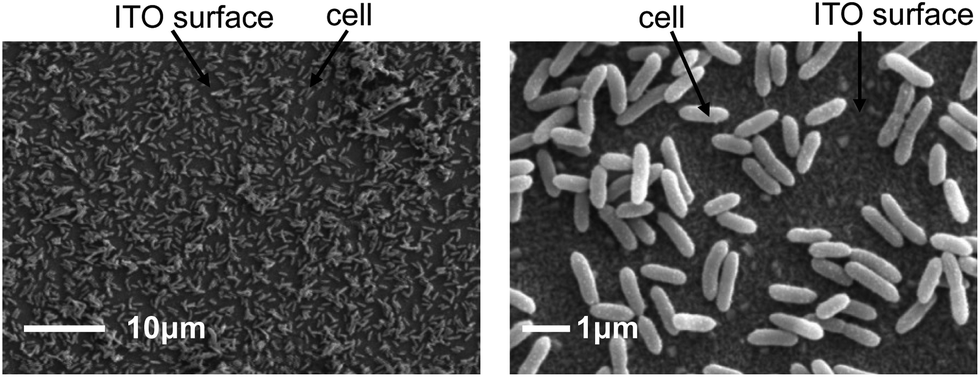 | ||
| Fig. 3 SEM images of the ITO electrode surface after 50 h of electrochemical cultivation of G. sulfurreducens at 0.4 V (vs. SHE) in the presence of 10 mM acetate. Filament-like structures were hardly detected. | ||
To examine the specific interactions between flavins and OM c-Cyts, we used a mutant strain (ΔomcBEST) of G. sulfurreducens lacking OmcB, OmcE, OmcS, and OmcT, which are the major multi-heme c-Cyts localized on the OM of Geobacter.19 When cultured at +0.4 V in the presence of 10 mM acetate, current production by the ΔomcBEST strain was highly impaired (Fig. S8†). In addition, the peak position and Ip of RF displayed a negative shift of 75 mV and 20-fold decrease, respectively, compared to those of the wild-type (WT) strain (Fig. 4a and S9†). Under FMN-supplemented conditions, the Ep of ΔomcBEST also exhibited a 50 mV negative shift in the DP voltammogram and a large Ip decrease compared to that of WT (Fig. S10†). These significant effects on Ep observed for the ΔomcBEST strain demonstrate that both FMN and RF associate with OmcB, OmcE, OmcS, or OmcT c-Cyt proteins to enhance the rate of EET in WT G. sulfurreducens cells. Such specific association of RF with OM c-Cyts of G. sulfurreducens cells was also confirmed by the fluorescence analysis of the OM fraction,20 where less flavin contents were observed for the OM fraction of ΔomcBEST cells compared with that of WT (Fig. S11†). However, in the system without omcBEST genes, the RF peak current at −0.125 V and Ic still showed a positive correlation with a comparable slope value to WT (Fig. S9c†). This result suggests that, in addition to omcBEST proteins, other types of OM c-Cyts in the Geobacter WT strain associate with RF to mediate EET (Fig. 4). This situation differs from the EET process reported for S. oneidensis MR-1, which is unable to bind FMN in the absence of a single binding protein (MtrC).14 This finding is also consistent with the wider variety of c-Cyts encoded in the genome of G. sulfurreducens compared to that of S. oneidensis MR-1.21 Together, these results confirm that RF and FMN associate with OM c-Cyts of G. sulfurreducens and serve as redox cofactors (Fig. 4), and that Geobacter is more flexible with respect to flavin uptake and binding mechanisms than S. oneidensis MR-1. In addition, because the peak current for bound-RF decreased when Ic was reduced by the addition of a metabolic inhibitor (Fig. 2d), reduced hemes in OM c-Cyts of G. sulfurreducens play an important role in the interaction with flavins, as has also been reported in MR-1.15
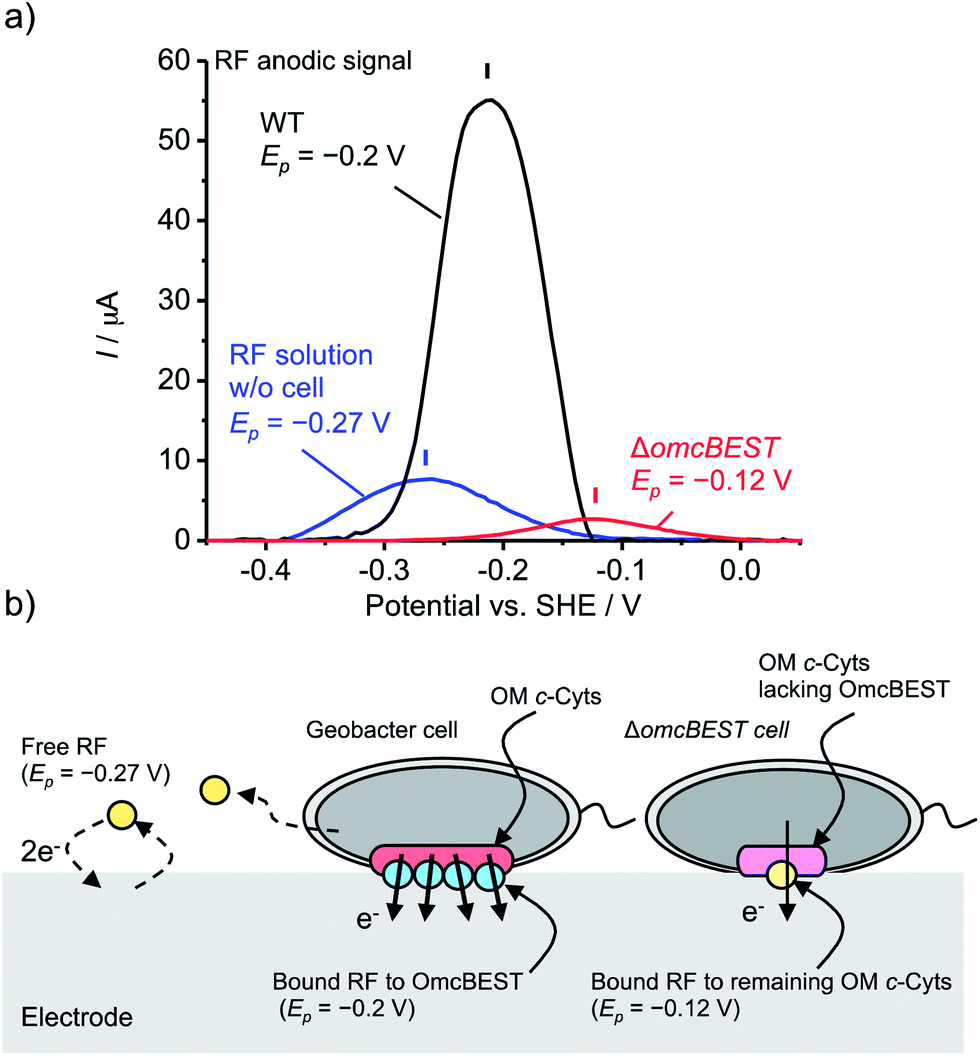 | ||
| Fig. 4 (a) Base-line subtracted differential pulse (DP) voltammograms of riboflavin (RF) for cell-free RF 10 μM solution (blue) and for G. sulfurreducens wild type (WT) (black) and ΔomcBEST strain (red) in the presence of 2 μM RF. (b) Schematic model illustration of interactions between RF and OM c-Cyts in extracellular electron transport (EET) processes. Free RF (yellow dots) scarcely involves in EET (Ep = −0.27 V vs. SHE), but bound RF associated with OM c-Cyts, omcBEST proteins (blue dots), works as a major EET pathway (Ep = −0.2 V). Although RF still binds to the remaining OM c-Cyts in the absence of omcBEST (Ep at −0.12 V) (a pale yellow dot), a decrease in the amount of bound RF results in a significantly lower current production compared with WT. | ||
Conclusions
We have provided the first experimental evidence for the involvement of self-secreted flavin in EET in G. sulfurreducens. As the flavin uptake mechanism in G. sulfurreducens appears to be similar to that of S. oneidensis MR-1,14 and that a wide range of microorganisms possesses homologous OM c-Cyt protein complexes to these two species,2,22 flavin may be a universal factor for efficient EET at bacteria/solid interfaces. In addition, if reduced hemes in OM c-Cyts govern the interaction with flavin, the intracellular redox homeostasis of Geobacter cells could be maintained by the release and binding of flavin, as is observed in MR-1,15 given that the oxidation state of hemes in OM c-Cyts reflects the balance between the electron input from respiration and the output by EET. Our present experiments were focused on monolayers of cells, where conductive filaments (nanowires) were not present; thus, it will be of great interest to examine the interaction of flavins with conductive filaments in thick biofilms, where these filaments may also play a role in intercellular EET. Recent theoretical analysis of charge flow along conductive filaments highlighted the importance of a multi-step hopping transport mechanism with charge localizing sites separated by less than 1 nm and reorganization energies lower than those known in biology.13 As arrays of OM c-Cyts such as OmcS expressed on the conductive filaments of G. sulfurreducens have been implicated as charge carriers,23 a specific flavin association with c-Cyts could also play an extensive role in multi-step charge hopping through thick biofilms. Further studies on the uptake of self-secreted flavins as the bound-cofactor of OM c-Cyts will serve for the mechanistic understanding of the complex EET reactions and also for optimization of microbial fuel cells6 and the bioremediation of waste waters and contaminated sediments.7Acknowledgements
We thank Prof. D. R. Lovley of the University of Massachusetts for kindly providing the ΔomcBEST mutant. This work was financially supported by a Grant-in-Aid for Specially Promoted Research of the Japan Society for Promotion of Science (JSPS) KAKENHI Grant Number 24000010.References
- K. H. Nealson and D. Saffarini, Annu. Rev. Microbiol., 1994, 48, 311 CrossRef CAS PubMed; D. R. Lovley, Annu. Rev. Microbiol., 2012, 66, 391 CrossRef PubMed.
- J. K. Fredrickson, M. F. Romine, A. S. Beliaev, J. M. Auchtung, M. E. Driscoll, T. S. Gardner, K. H. Nealson, A. L. Osterman, G. Pinchuk, J. L. Reed, D. A. Rodionov, J. L. M. Rodrigues, D. A. Saffarini, M. H. Serres, A. M. Spormann, I. B. Zhulin and J. M. Tiedje, Nat. Rev. Microbiol., 2008, 6, 592 CrossRef CAS PubMed.
- S. Matsuda, H. Liu, S. Kato, K. Hashimoto and S. Nakanishi, Environ. Sci. Technol., 2011, 45, 10163 CrossRef CAS PubMed; H. W. Harris, M. Y. El-Naggar, O. Bretschger, M. J. Ward, M. F. Romine, A. Y. Obraztsova and K. H. Nealson, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 326 CrossRef PubMed.
- Z. M. Summers, H. E. Fogarty, C. Leang, A. E. Franks, N. S. Malvankar and D. R. Lovley, Science, 2010, 330, 1413 CrossRef CAS PubMed; R. Nakamura, F. Kai, A. Okamoto, G. J. Newton and K. Hashimoto, Angew. Chem., Int. Ed., 2009, 48, 508 CrossRef PubMed.
- S. Kato, K. Hashimoto and K. Watanabe, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 10042 CrossRef CAS PubMed.
- D. R. Lovley, Nat. Rev. Microbiol., 2006, 4, 497 CrossRef CAS PubMed; U. Schroder, J. Niessen and F. Scholz, Angew. Chem., Int. Ed., 2003, 42, 2880 CrossRef PubMed.
- B. K. Amos, Y. Sung, K. E. Fletcher, T. J. Gentry, W. M. Wu, C. S. Criddle, J. Zhou and F. E. Loffler, Appl. Environ. Microbiol., 2007, 73, 6898 CrossRef CAS PubMed.
- H. T. Dinh, J. Kuever, M. Mussmann, A. W. Hassel, M. Stratmann and F. Widdel, Nature, 2004, 427, 829 CrossRef CAS PubMed.
- D. K. Newman and R. Kolter, Nature, 2000, 405, 94 CrossRef CAS PubMed.
- E. Marsili, D. B. Baron, I. D. Shikhare, D. Coursolle, J. A. Gralnick and D. R. Bond, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 3968 CrossRef CAS PubMed.
- T. A. Clarke, M. J. Edwards, A. J. Gates, A. Hall, G. F. White, J. Bradley, C. L. Reardon, L. Shi, A. S. Beliaev, M. J. Marshall, Z. M. Wang, N. J. Watmough, J. K. Fredrickson, J. M. Zachara, J. N. Butt and D. J. Richardson, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 9384 CrossRef CAS PubMed; D. J. Richardson, J. N. Butt and T. A. Clarke, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 7537 CrossRef PubMed; H. Liu, G. J. Newton, R. Nakamura, K. Hashimoto and S. Nakanishi, Angew. Chem., Int. Ed., 2010, 49, 6596 CrossRef PubMed.
- N. S. Malvankar, M. Vargas, K. P. Nevin, A. E. Franks, C. Leang, B.-C. Kim, K. Inoue, T. Mester, S. F. Covalla, J. P. Johnson, V. M. Rotello, M. T. Tuominen and D. R. Lovley, Nat. Nanotechnol., 2011, 6, 573 CrossRef CAS PubMed; Y. A. Gorby, S. Yanina, J. S. McLean, K. M. Rosso, D. Moyles, A. Dohnalkova, T. J. Beveridge, I. S. Chang, B. H. Kim, K. S. Kim, D. E. Culley, S. B. Reed, M. F. Romine, D. A. Saffarini, E. A. Hill, L. Shi, D. A. Elias, D. W. Kennedy, G. Pinchuk, K. Watanabe, S. Ishii, B. Logan, K. H. Nealson and J. K. Fredrickson, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 11358 CrossRef PubMed.
- N. F. Polizzi, S. S. Skourtis and D. N. Beratan, Faraday Discuss., 2012, 155, 43 RSC.
- D. R. Bond and D. R. Lovley, Appl. Environ. Microbiol., 2003, 69, 1548 CrossRef CAS.
- A. Okamoto, K. Hashimoto, K. H. Nealson and R. Nakamura, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 7856 CrossRef CAS PubMed.
- N. J. Kotloski and J. A. Gralnick, mBio, 2013, 4, 1 CrossRef PubMed.
- H. von Canstein, J. Ogawa, S. Shimizu and J. R. Lloyd, Appl. Environ. Microbiol., 2008, 74, 615 CrossRef CAS PubMed.
- G. Reguera, K. P. Nevin, J. S. Nicoll, S. F. Covalla, T. L. Woodard and D. R. Lovley, Appl. Environ. Microbiol., 2006, 72, 7345 CrossRef CAS PubMed.
- K. Inoue, C. Leang, A. E. Franks, T. L. Woodard, K. P. Nevin and D. R. Lovley, Environ. Microbiol. Rep., 2011, 3, 211 CrossRef CAS PubMed; J. W. Voordeckers, B. C. Kim, M. Izallalen and D. R. Lovley, Appl. Environ. Microbiol., 2010, 76, 2371 CrossRef PubMed; C. Leang, X. Qian, T. Mester and D. R. Lovley, Appl. Environ. Microbiol., 2010, 76, 4080 CrossRef PubMed.
- K. Inoue, X. L. Qian, L. Morgado, B. C. Kim, T. Mester, M. Izallalen, C. A. Salgueiro and D. R. Lovley, Appl. Environ. Microbiol., 2010, 76, 3999 CrossRef CAS PubMed.
- B. A. Methe, K. E. Nelson, J. A. Eisen, I. T. Paulsen, W. Nelson, J. F. Heidelberg, D. Wu, M. Wu, N. Ward, M. J. Beanan, R. J. Dodson, R. Madupu, L. M. Brinkac, S. C. Daugherty, R. T. DeBoy, A. S. Durkin, M. Gwinn, J. F. Kolonay, S. A. Sullivan, D. H. Haft, J. Selengut, T. M. Davidsen, N. Zafar, O. White, B. Tran, C. Romero, H. A. Forberger, J. Weidman, H. Khouri, T. V. Feldblyum, T. R. Utterback, S. E. Van Aken, D. R. Lovley and C. M. Fraser, Science, 2003, 302, 1967 CrossRef CAS PubMed; J. F. Heidelberg, I. T. Paulsen, K. E. Nelson, E. J. Gaidos, W. C. Nelson, T. D. Read, J. A. Eisen, R. Seshadri, N. Ward, B. Methe, R. A. Clayton, T. Meyer, A. Tsapin, J. Scott, M. Beanan, L. Brinkac, S. Daugherty, R. T. DeBoy, R. J. Dodson, A. S. Durkin, D. H. Haft, J. F. Kolonay, R. Madupu, J. D. Peterson, L. A. Umayam, O. White, A. M. Wolf, J. Vamathevan, J. Weidman, M. Impraim, K. Lee, K. Berry, C. Lee, J. Mueller, H. Khouri, J. Gill, T. R. Utterback, L. A. McDonald, T. V. Feldblyum, H. O. Smith, J. C. Venter, K. H. Nealson and C. M. Fraser, Nat. Biotechnol., 2002, 20, 1118 CrossRef PubMed.
- J. E. Butler, N. D. Young and D. R. Lovley, BMC Genomics, 2009, 10, 103 CrossRef PubMed.
- C. Leang, X. L. Qian, T. Mester and D. R. Lovley, Appl. Environ. Microbiol., 2010, 76, 4080 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Figures and methods. See DOI: 10.1039/c3ee43674h |
| This journal is © The Royal Society of Chemistry 2014 |