A dialkynoyl analogue of DOPE improves gene transfer of lower-charged, cationic lipoplexes†
Steven Fletcher‡a, Ayesha Ahmada, Eric Perouzelab, Michael R. Jorgensen*ab and Andrew D. Miller*ab
aImperial College Genetic Therapies Centre, Department of Chemistry, Flowers Building, Armstrong Road, Imperial College London, London, UK SW7 2AZ. E-mail: a.miller@imperial.ac.uk; Fax: +44-(0)20-7594-5803; Tel: +44-(0)20-7594-5869
bIC-Vec Ltd, 13 Prince's Gardens, London, UK SW7 1NA. E-mail: m.jorgensen@icvec.com; Fax: +44-(0)20-7594-1061; Tel: +44-(0)20-7594-3150
First published on 6th December 2005
Abstract
Positively-charged gene delivery agents, such as cationic liposomes, typically prepared by mixing a cationic lipid and a neutral lipid in a 1 : 1 molar ratio, exhibit a fundamental flaw: on the one hand, the charge encourages cell uptake; on the other hand, the charge leads to aggregation in vivo with anionic serum components. We herein report a more phase-stable analogue of the zwitterionic and fusogenic lipid DOPE that allows for the reduction of the cationic lipid component of the liposome from 50 to 9 mol% with almost no apparent loss in transfection activity. This reduction in charge may induce important in vivo stability whilst still imparting high cell uptake and transgene expression.
Cationic liposomes for non-viral gene therapy1 are typically prepared by mixing a cationic lipid with a neutral, helper lipid, such as dioleoylphosphatidylethanolamine (DOPE (1)).2 The cationic lipid serves two purposes: complexation with anionic DNA and interaction with anionic cellular membranes (induction of endocytosis and endosomolysis), whereas the neutral, helper lipid alters the physical properties of the delivery vehicle so as to adjust its functionality. In particular, helper lipids are believed to assist in endosomal escape. DOPE (1), one of the more commonly used neutral lipids, exhibits lipid polymorphism under various concentration and temperature conditions. In the presence of cationic lipids, DOPE (1)-containing liposomes may be organised into either the stable, bilayer (Lα) phase or the fusogenic, inverted hexagonal, (HII) phase and are found to greatly enhance transfection efficiency (gene delivery and expression) in vitro.3–7 This is due to the overwhelming preference of DOPE (1) for the fusogenic HII phase at 37 °C, promoting fusion with cellular membranes,8,9 especially the endosomal membrane, thereby facilitating escape of the genetic material from the digestive, endocytic cycle.
However, due to the low Lα/HII phase transition temperature (Th = 10 °C),10 DOPE (1) has limited utility in vivo because the very feature of DOPE (1) that promotes transfection in vitro renders cationic liposomes too unstable in the more complex and hazardous in vivo environment.11 Moreover, it is desirable to reduce the magnitude of charge in cationic liposomes, since this charge leads to rapid liposome clearance by the reticuloendothelial system (RES) in vivo.12 Replacement of the cationic lipid with increasing amounts of DOPE (1) leads to a gradual loss of transfection in vitro and an even more substantial loss in vivo, due to less cationic charge and increased liposome structural instability.13,14 Furthermore, replacement of the cationic lipid with a bilayer-forming lipid likewise leads to a loss in transfection,5,6 due to excessive stability resulting from the lack of a fusogenic component. For sustained transfection, it is advantageous that the neutral co-lipid should be able to access the fusogenic HII phase under physiological conditions, such that there is a “trade-off” where loss of transfection potency due to reduced cationic lipid is compensated for by the inclusion of increased fusogenic lipid. In order to prepare lower-charged, yet highly-transfecting cationic liposomes, one approach might be to create analogues of DOPE (1) in which the Lα/HII phase transition temperature is nearer the physiological temperature of 37 °C.
In this manuscript, we report the synthesis of a series of dialkynoyl analogues of DOPE (1). We hypothesised that the inherent instability (HII phase) of DOPE (1) may be suitably altered by substituting the cis-double bond in the oleoyl fatty acid chains with an acetylenic bond, in varying locations along the chains, without impairing transfection. By replacing the cis-double bonds with the less “kinked” triple bonds, it was anticipated that the intermolecular packing would be improved, such that the Lα/HII phase transition would concomitantly be raised, affording DOPE (1)-analogues that would be more phase stable under laboratory preparation conditions, yet still allow access to the fusogenic HII-phase under physiological conditions. Moreover, we considered that such harnessed fusogenicity would allow for the incorporation of reduced amounts of cationic lipid without impairing transfection, thereby promoting such systems for in vivo applications as well.
Fig. 1 shows the modifications that we made to the DOPE (1) molecule, replacing the cis-double bonds at position 9 of the fatty acid chains with triple bonds at positions 4, 9 and 14, as the isomeric compounds 2, 3 and 4, respectively. As a representative example, the synthesis of 3 is given in Scheme 1. First, the hydroxyl group of 8-bromooctan-1-ol (5) was protected quantitatively as its base-stable THP ether, and, in order to render the resultant bromo ether more reactive for the subsequent transformation, Br was substituted for I under Finkelstein conditions, to give 6. Treatment of 1-decyne/n-BuLi with 6 at 0 °C then furnished internal alkyne 7 in good yield (72%). THP removal followed by oxidation of the liberated primary alcohol to acid 9 (63% for the two steps) generated the moiety required to conjugate to the zwitterionic phospholipid headgroup 10. Activation of acid 9 with CDI allowed the coupling to glycerophosphocholine (10) to proceed smoothly and in good yield (67%), to afford di(octadec-9-ynoyl)phosphatidylcholine (DS(9-yne)PC, 11). Subsequent enzymatic transphosphatidylation15 of PC 11 with phospholipase D in the presence of ethanolamine furnished di(octadec-9-ynoyl)phosphatidylethanolamine (DS(9-yne)PE, 3) in an excellent yield of 94%. The other lipids, DS(4-yne)PE (2) and DS(14-yne)PE (4), were prepared in similar fashions and in similar yields (refer to the electronic supplementary information).
 | ||
| Fig. 1 DOPE (1) and its dialkynoyl analogues (2–4). | ||
 | ||
| Scheme 1 (a) DHP, PPTS, CH2Cl2, 0 °C → RT, 16 h, 93%; (b) NaI, acetone, Δ, 15 min, 89%; (c) (i) 1-decyne, n-BuLi, THF, 0 °C, 15 min; (ii) HMPA, 0 °C, 15 min; (iii) 6, THF, 0 °C → RT; 48 h, 72%; (d) p-TsOH, MeOH, RT, 20 h, 92%; (e) Jones's reagent, 0 °C, 2 h, 69%; (f) (i) CDI, CHCl3, RT, 30 min; (ii) 10, DBU, DMSO, RT, 7 h, 67%; (g) phospholipase D, ethanolamine, NaOAc–CaCl2 buffer, pH 6.5, CHCl3, 30 °C, 94%. | ||
Next, the Lα/HII phase transition temperatures (Th) of these dialkynoyl analogues of DOPE (1) were determined by 31P NMR spectroscopy in neutral D2O.16 A typical bilayer (Lα) lipid, such as dioleoylphosphatidylcholine (DOPC—the choline analogue of DOPE (1)), gives an asymmetric 31P NMR spectrum comprising a low field shoulder and a high field peak. Conversely, HII-preferring lipids, such as DOPE (1), give a lineshape with a reversed asymmetry.16 Hence, using the technique of 31P NMR, the Lα/HII phase transition temperatures were determined and are given in Table 1 (31P NMR data were corroborated by preliminary X-ray diffraction studies (data not shown)). As indicated, DS(4-yne)PE (2) exhibited the Lα phase up to 40 °C and beyond, and was therefore anticipated to be inhibitory towards transfection. DS(14-yne)PE (4) was considered to be potentially less inhibitory since this lipid appeared to co-exist in both the stable Lα and fusogenic HII phases between 30 and 55 °C. By contrast, DS(9-yne)PE (3) exhibited the HII phase at physiological temperatures. The 31P NMR static powder pattern spectra (Fig. 2) of DS(9-yne)PE (3) taken at 5 °C intervals from 20 to 40 °C show a smooth transition from the stable Lα phase to the fusogenic HII phase at around 30 °C. This is approximately a 20 °C rise in the Th, relative to DOPE (1),10 and it is likely that this change in Th is due to the existence of tighter intermolecular packing in DS(9-yne)PE (3), as predicted. Moreover, this Th was considered ideal for our purposes, since it allows for the preparation of stable liposomes (Lα phase) under laboratory conditions (25 °C), yet also allows access to the fusogenic HII phase under physiological conditions (37 °C).
| PE lipid | Lα/HII phase transition temperature (°C) |
|---|---|
| DOPE (1) | 8-1716 |
| DS(4-yne)PE (2) | > 40 |
| DS(9-yne)PE (3) | 25–35 |
| DS(14-yne)PE (4) | 30–55 |
 | ||
| Fig. 2 31P NMR static powder pattern spectra of DS(9-yne)PE (3) obtained over a range of temperatures. The Lα/HII phase transition occurs around 25–35 °C. | ||
In an effort to correlate the structural properties of the lipids to their biofunctionalities, in vitro testing on a Panc-1 cell line was performed. Since cationic charge is needed for DNA-complexation and cellular uptake, each of the four PE lipids were mixed in various molar ratios with the potent, tri-amino cationic lipid N1-cholesteryloxycarbonyl-3,7-diazanonane-1,3-diamine (CDAN,1712, Fig. 3) to generate cationic liposomes that were then complexed with 0.5 µg of plasmid DNA harbouring the luciferase reporter gene, in a 12 : 1, w/w ratio. This thereby afforded the transfection-competent cationic liposome–DNA particles (lipoplex, LD), exhibiting a range of N : P (number of protonatable amines : number of phosphate negative charges) ratios, as detailed in Table 2. As indicated, the N : P ratios decrease as less cationic lipid is incorporated into the LD particle. Whilst reduced cationic charge is anticipated to cause a drop in transfection potency through reduced cellular uptake, the purpose of our study is to compensate for this by incorporating increasing fusogenic lipid to assist in endosomal escape, which is another barrier to the transfection process. Panc-1 cells in 100% DMEM, seeded in 24-well plates, were then treated with these LD systems for 5 h, after which time, the cells were washed with PBS buffer, incubated with DMEM supplemented with 10% fetal calf serum, for a further 24 h, then harvested. Transfection results were determined as relative light units (RLU) of luciferase standardised by total cell protein (BCA assay).
| CDAN : PEb | CDAN : DNAc | N : P ratioa |
|---|---|---|
| a N : P ratios were calculated using the following formula: (MW_DNA/MW CDAN) × (no. of amines_CDAN/no. of phosphates_DNA) × weight ratio.b Ratio is mol : mol.c Ratio is weight : weight. | ||
| 1 : 1 | 5.57 : 1 | 8.35 : 1 |
| 1 : 4 | 2.17 : 1 | 3.26 : 1 |
| 1 : 6 | 1.55 : 1 | 2.32 : 1 |
| 1 : 8 | 1.20 : 1 | 1.80 : 1 |
| 1 : 10 | 0.98 : 1 | 1.47 : 1 |
 | ||
| Fig. 3 N1-Cholesteryloxycarbonyl-3,7-diazanonane-1,3-diamine (CDAN, 12). | ||
Transfection data for CDAN : PE LD systems, at reducing molar ratios of cationic lipid to neutral lipid are shown in Fig. 4. Due to increasing content (to as much as 91 mol%) of PE lipid in the cationic liposome and LD formulations, it is reasonable to draw correlations between the phase behaviour of the PE lipids in isolation and the transfection properties of their corresponding LD particles. Whilst it is acknowledged that DNA complexation may also alter the fluidity of the liposome bilayer, this work focuses on the fluidity of liposome bilayers as dictated by lipid stability. For CDAN : DOPE (1) LD particles, the transfection efficiency was found to decrease systematically as the ratio was changed from 1 : 1 to 1 : 10. This is to be expected, since the magnitude of cationic charge was being systematically reduced and the highly-fusogenic lipid content was being increased. It is likely that DOPE (1), whose Th = 10 °C, rendering the resultant LD particles too fusogenic, such that these particles may have destabilised outside of the cell. A similar rationale may explain the extremely poor transfection activity of DOPE (1)-containing liposomes in vivo.11 DS(4-yne)PE (2) showed a similar reduction in transfection, although we consider that this was due to the enhanced stability associated with the lamellar phase, as exhibited by this PE lipid at 37 °C. Whilst DS(14-yne)PE (4) predominantly exhibits the stable Lα phase, 31P NMR data suggests there is also limited HII phase behaviour at 37 °C, and this may account for the less-marked drop in transfection down to the ratio of 1 : 8, CDAN : DS(4-yne)PE (2). It is noteworthy that DS(14-yne)PE (4) outperforms DS(4-yne)PE (2) by approximately an order of magnitude at the CDAN : PE ratios of 1 : 6 and 1 : 8; it is likely that this is due to the different polymorphic phase behaviours of these lipids. The similarity in transfection levels at the CDAN : PE ratio of 1 : 10 for DOPE (1), DS(4-yne)PE (2) and DS(14-yne)PE (4) may be a consequence of excessive instability (in the case of 1) or excessive stability (in the cases of 2 and 4), such that the cationic charge of CDAN alone governed transfection efficiency. Most interestingly, the transfection potency of CDAN : DS(9-yne)PE (3)-based LD particles was maintained across the series from 1 : 1 to 1 : 10, and zeta potential measurements confirmed there was a reduction in charge of CDAN : DS(9-yne)PE (3) liposomes 1 : 1 to 1 : 10 of +93.4 ± 3.4 to +62.4 ± 0.7 mV. This suggests that the reduction of cationic charge resulting from the incorporation of less cationic lipid (CDAN) is indeed offset by the incorporation of the fusogenic lipid DS(9-yne)PE (3), whose Th has been fine-tuned to a more physiologically-relevant temperature. It is remarkable to note that the simple replacement of the cis-double bonds of DOPE with triple bonds in the same positions leads to an increase in transfection potency of almost two orders of magnitude, when comparing the DOPE (1) and DS(9-yne)PE (3) transfection data for the ratio of 1 : 10, CDAN : PE.
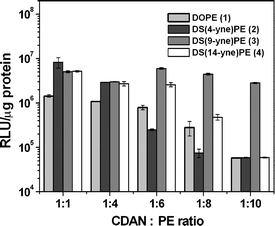 | ||
| Fig. 4 Transfection activities of lipoplexes (12 : 1, w/w, lipid : DNA) prepared from cationic liposomes comprising different ratios of CDAN : PE. Data are expressed as relative light units (RLU) of reporter gene luciferase, standardised by total cell protein. | ||
In conclusion, the substitution of the cis-double bonds of DOPE (1) with triple bonds in varying positions along the fatty acid chains raises the Lα/HII phase transition temperature, thereby affording more phase stable lipids. By 31P NMR spectroscopy, we have shown that DS(9-yne)PE (3) has an Lα/HII phase transition temperature of approximately 30 °C and allows for the preparation of lower-charged, cationic liposomes (and LD particles) without imparing transfection since the fusogenic HII phase is accessed at 37 °C. Currently, we are performing further transfection studies in the presence of serum to begin to emulate the in vivo environment. In conjunction with toxicity and X-ray diffraction studies we hope to elucidate further the promising behaviour of these PE-analogues. These results shall be reported in due course.
Acknowledgements
Dr Steven Fletcher gratefully acknowledges funding from Mitsubishi Chemicals Corporation and IC-Vec, Ltd. Funding for Dr Ayesha Ahmad provided by National Science Foundation, USA, IRFP grant. We thank James Clarke and Dr Rob Law for assistance in running the 31P NMR experiments.References
- P. L. Felgner, T. R. Gadek, M. Holm, R. Roman, H. W. Chan, M. Wenz, J. P. Northrop, G. M. Ringold and M. Danielsen, Proc. Natl. Acad. Sci. U. S. A., 1987, 84, 7413–7417.
- A. D. Miller, Angew. Chem., Int. Ed., 1998, 37, 1768–1785 CrossRef.
- J. Y. Legendre and F. C. Szoka, Jr., Proc. Natl. Acad. Sci. U. S. A., 1993, 90(3), 893–7 CAS.
- J. H. Felgner, R. Kumar, C. N. Sridhar, C. J. Wheeler, Y. J. Tsai, R. Border, P. Ramsey, M. Martin and P. L. Felgner, J. Biol. Chem., 1994, 269, 2550–2561 CAS.
- H. Farhood, N. Serbina and L. Huang, Biochim. Biophys. Acta, 1995, 1235, 289–295 CrossRef CAS.
- I. Koltover, T. Salditt, J. O. Radler and C. R. Safinya, Science, 1998, 281, 78–81 CrossRef CAS.
- R. Koynova, H. S. Rosenzweig, L. Wang, M. Wasielewski and R. C. MacDonald, Chem. Phys. Lipids, 2004, 129(2), 183–94 CrossRef CAS.
- D. C. Litzinger and L. Huang, Biochim. Biophys. Acta, 1992, 1113, 201–227 CAS.
- S. M. Gruner, P. R. Cullis, M. J. Hope and C. P. Tilcock, Annu. Rev. Biophys. Biophys. Chem., 1985, 14, 211–38 CrossRef CAS.
- R. N. Lewis, D. A. Mannock, R. N. McElhaney, D. C. Turner and S. M. Gruner, Biochemistry, 1989, 28(2), 541–8 CrossRef CAS.
- S. Li, M. A. Rizzo, S. Bhattacharya and L. Huang, Gene Ther., 1998, 5, 930–937 CrossRef CAS.
- J. Senior, J. C. Crawley and G. Gregoriadis, Biochim. Biophys. Acta, 1985, 839(1), 1–8 CrossRef CAS.
- R. J. Lee and L. Huang, Crit. Rev. Ther. Drug Carrier Syst., 1997, 14(2), 173–206 Search PubMed.
- A. D. Miller, Curr. Med. Chem., 2003, 10(14), 1195–211 CrossRef CAS.
- P. Wang, M. Schuster, Y.-F. Wang and C.-H. Wong, J. Am. Chem. Soc., 1993, 115, 10487–10491 CrossRef CAS.
- P. R. Cullis and B. de Kruijff, Biochim. Biophys. Acta, 1978, 513(1), 31–42 CAS.
- R. G. Cooper, C. J. Etheridge, L. Stewart, J. Marshall, S. Rudginsky, S. H. Cheng and A. D. Miller, Chem. Eur. J., 1998, 4, 137–151 CrossRef.
Footnotes |
| † Electronic supplementary information (ESI) available: Experimental details, 1H and 13C NMR, mass spectral analysis and HPLC retention times. See DOI: 10.1039/b514532e |
| ‡ Current address: Department of Chemistry, 225, Prospect Street, Yale University, P.O. Box 208107, New Haven, CT 06511-8107, USA. |
| This journal is © The Royal Society of Chemistry 2006 |