
Efficient transcription initiation in bacteria: an interplay of protein–DNA interaction parameters†
Abstract
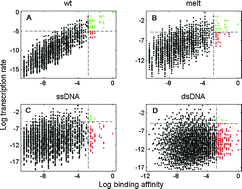
As the first, and usually rate-limiting, step of transcription initiation, bacterial RNA polymerase (RNAP) binds to double stranded DNA (dsDNA) and subsequently opens the two strands of DNA (the open complex formation). The rate determining step in the open complex formation is opening of a short (6 bp) DNA called the −10 region, which interacts with RNAP in both dsDNA and single stranded (ssDNA) forms. Accordingly, formation of the open complex depends on (physically independent) domains of RNAP that interact with ssDNA and dsDNA, as well as on parameters of DNA melting and sequences of −10 regions. We here aim to understand how these different interactions are mutually related to ensure efficient open complex formation. To achieve this, we use a recently developed biophysical model of transcription initiation, which allows the calculation of the kinetic parameters of transcription initiation on the scale of whole genome. We consequently investigate kinetic properties of sequences derived from all E. coli intergenic regions, and from more than 300 experimentally confirmed E. coli σ70 promoters. We find that interaction specificities of σ70 DNA binding domains reduce the number of sequences where RNAP binds strongly, but forms the open complex too slowly to achieve functional transcription (so-called poised promoters). However, we find that, despite this reduction, there is still a significant number of such poised promoters in the intergenic regions, which may provide a major source of false positives in genome-wide searches of transcription start sites. Furthermore, we surprisingly find that sequences of −10 regions of the functional promoters increase the extent of RNAP poising, which we interpret in terms of an extension of a recently proposed model of promoter recognition (‘mix-and-match model’) to kinetic parameters. Overall, our results allow better understanding of the design of σ70 DNA binding domains and promoter sequences, and place a fundamental limit on accuracy of methods for promoter detection that are based on strong RNAP binding (e.g. ChIP-chip).
 Please wait while we load your content...
Please wait while we load your content...