
Biphasic modulation of tau liquid–liquid phase separation by polyphenols†
Hariharan
Moorthy
 ,
Nimsha
Kamala
,
Madhu
Ramesh
and
Thimmaiah
Govindaraju
*
,
Nimsha
Kamala
,
Madhu
Ramesh
and
Thimmaiah
Govindaraju
*
Bioorganic Chemistry Laboratory, New Chemistry Unit, Jawaharlal Nehru Centre for Advanced Scientific Research (JNCASR), Bengaluru 560064, Karnataka, India. E-mail: tgraju@jncasr.ac.in
First published on 22nd March 2024
Abstract
Molecular tools that modulate tau liquid–liquid phase separation (LLPS) promise to treat tauopathies. We screened a set of polyphenols and demonstrated concentration-dependent biphasic modulation of tau LLPS by gallic acid (GA), showcasing its ability to expedite the liquid-to-gel transition in tau condensates and effectively impede the formation of deleterious fibrillar aggregates.
Tauopathies including Alzheimer's disease (AD) are marked by the abnormal misfolding and pathological accumulation of intrinsically disordered protein tau.1–3 Tau controls the dynamics of microtubules and influences the activity of motor proteins.4,5 Under pathological conditions, tau undergoes unusual post-translational modifications and aggregations, leading to toxic neurofibrillary tangles (NFTs).4,6,7 Natural phenols, including (−)-epigallocatechin-3-gallate (EGCG), tannic acid (TA), myricetin, oleocanthal, curcumin, and rosmarinic acid have been shown to inhibit toxic tau aggregation.8–10 Thiophene-based small molecules that modulate both amyloid-β (Aβ) and tau aggregation and effectively protect neuronal cells have been identified.11 Methylene blue (MB) and leuco-methylthioninium (LMTM), bis(hydromethanesulphonate) were found to inhibit tau aggregation, however, their poor results in phase III clinical trials underscore the need for further investigation into tau aggregation mechanisms for the development of effective tau-targeted therapeutics.12,13
LLPS is a biological process where specific proteins segregate and form membrane-less dynamic organelles or biomolecular condensates.14 LLPS regulates cell signalling and stress response, but under pathology, they can lead to disease-associated aggregates.15–18 It is influenced by factors like concentration, pH, salt, RNA, and PTMs.16,19 Low RNA concentrations promote protein LLPS via constructive electrostatic interactions, while higher concentrations induce decondensation via long-range electrostatic repulsions.20 This biphasic behaviour is reminiscent of the regulatory dynamics observed in cellular protein/RNA condensates.16,21 Recent studies indicate tau readily undergoes LLPS to form liquid-like condensates.22,23 While phase-separated tau was found to nucleate microtubule bundles, its aberrant phase transitions increased local concentration, driving toxic aggregates.20,24 Identification of molecular modulators of LLPS that can disrupt aberrant phase transition, while preserving the functionality of LLPS is critical for devising new therapeutic strategies.25 In this regard, myricetin was found to slow the LLPS of tau and inhibit its aggregation.26 EGCG was shown to promote tau LLPS mainly by binding to its proline-rich domain.9 Suramin was found to induce tau LLPS to reduce the formation of heparin-induced tau aggregates.27 We have developed a cyclic dipeptide derivative that effectively inhibits and dissolves Zn-mediated tau condensates preventing the tau condensate-to-fibril transition.28 MB was shown to promote tau LLPS and accelerate the gel transition from matured tau droplets, thereby reducing the fibrillation and cytotoxicity.29
The complex role of LLPS in driving the fibrillation process necessitates the need for identifying molecular modulators of tau LLPS. In this study, we evaluated a set of simplest polyphenols (SPs) including 3,4 dihydroxy benzaldehyde (DHBA), GA, syringic acid (SA), 2,2-dithiosalicylic acid (DTSA), and TA for their ability to alter tau LLPS. Our findings reveal that GA and similar polyphenols including DHBA, SA, dopamine (DP), L-Dopa (LD) display biphasic modulation of tau LLPS. At lower concentrations, GA enhances tau LLPS, whereas at higher concentrations, it disrupts LLPS through a reentrant phase transition. GA promotes tau LLPS through a combination of hydrophobic and electrostatic interactions and expedites the liquid-to-gel transition of the tau condensates. Surprisingly, GA inhibits arachidonic acid (AA)-induced tau aggregation, thereby protecting neuronal cells from tau toxicity. These results not only establish the ability of polyphenols to control tau LLPS but also lay the groundwork for identifying compounds that could potentially regulate tau LLPS for therapeutic targeting of tauopathies.
The increase in turbidity and DN of tau in LLPS buffer (10 mM HEPES, 10 mM NaCl, 10% PEG, pH 7.4) compared to HEPES buffer (10 mM, pH 7.4) indicates phase separation (Fig. S1A–C, ESI†). Further, fluorescent-tagged tau incubated in LLPS buffer revealed fluorescent droplets of tau corroborating the above assay (Fig. S1D, ESI†). Next, SPs were evaluated for their ability to modulate the LLPS of tau (Fig. 1A). To investigate the influence of SPs on tau LLPS, we incubated full-length wild type 2N4R (WT) tau (10 μM) independently and in the presence of various SPs (20 μM) for 30 min. Turbidity increase upon incubation with GA, SA, DHBA and TA indicates possible phase separation (Fig. 1B).9 To confirm, DIC imaging was conducted after incubating tau (5 μM) with SP (10 μM) for 30 minutes (Fig. 1C and Fig. S2, ESI†). Quantitative analysis confirmed increased tau DN in the presence of SPs, with the highest observed for GA (Fig. 1C and D). Tau undergoes LLPS with crowding agents like PEG or dextran. Notably, GA induces robust condensation of tau in the absence of any crowding agent. Pyrogallol (PG), structurally similar to GA, could not modulate tau LLPS, emphasizing the intrinsic role of molecular variations in LLPS modulation.9,30,31 Furthermore, a GA dose-dependent turbidity assay and DIC imaging indicated a biphasic nature (Fig. 2A). Tau LLPS was augmented across a GA concentration range of 1–10 μM, as evidenced by the increasing DN (Fig. 2B and C). However, a significant decrease in DN was observed at concentrations of 30–50 μM, suggesting a reentrant phase transition. Lower concentrations of GA promote tau LLPS through attractive forces among tau molecules, while higher concentrations induce decondensation via repulsive GA–tau interactions. Subsequent TEM studies showed that lower concentrations of GA facilitated droplet formation (Fig. 2D). However, with increasing GA concentration, we observed droplet coalescence (see inset) into amorphous aggregates, altering the fate of the metastable state and underlining the biphasic behaviour of GA in tau LLPS (Fig. 2D). Intrigued by the higher number of droplets at 10 μM of GA, we performed a time dependent DIC and AFM which revealed that the droplets were relatively stable over a period of 36 h (Fig. 2E, F and Fig. S4 and S5, ESI†). Next, the impact of pH on GA-induced tau LLPS showed a significant increase in DN at pH 5.2, comparable levels at pH 3.2 and 7.4, and a drastic decrease in DN at pH 9.2 (Fig. S6, ESI†). Tau coacervates via electrostatic interactions, forming either simple coacervates with its charged segments or complex coacervates with polyanions.20,29 DIC imaging quantification showed that WT tau LLPS was disrupted by NaCl, but not by 1,6 hexanediol (HD) or dimethylformamide (DMF) confirming simple coacervation primarily depends on electrostatic interactions (Fig. S7A and B, ESI†).9,22,27 However, the GA-induced tau condensates were sensitive to both HD and NaCl, but not to DMF (Fig. 3A and Fig. S8, ESI†). This markedly different response indicated that GA promotes tau LLPS through hydrophobic and electrostatic interactions, similar to MB-promoted LLPS.29 Additionally, it is important to note that GA augments tau LLPS and modulates its intermolecular interactions, possibly modulating the metastable state of tau LLPS. From a thermodynamic standpoint, tau LLPS formation is energetically favorable.32 In GA-induced LLPS, hydrophobic and electrostatic interactions suggest a propensity towards lower critical solution temperature (LCST), primarily driven by entropy.32,33 For such systems, peptide-bound water release compensates for entropy loss due to reduced peptide conformations during LLPS.32,34 The metastable LLPS state is influenced by GA, favoring thermodynamically lower free energy and potentially directing it towards a non-toxic pathway. This phenomenon leads to droplet coalescence and the formation of amorphous aggregates, elucidating the observed biphasic nature of tau LLPS. Next, we investigated the influence of GA on LLPS in other proteins like bovine serum albumin (BSA) and α-synuclein. Notably, no phase-separated droplets were detected with BSA (Fig. S9, ESI†). Intriguingly, we observed biphasic LLPS in α-synuclein, albeit at lower DN, across varying concentrations of GA. (Fig. S10, ESI†). Next, we used fluorescent-tagged tau to visualize tau droplets under confocal microscopy, revealing that GA (10 μM) enhances tau (5 μM) condensation. (Fig. S11, ESI†). Subsequent FRAP experiments demonstrated approximately 45% recovery with a half-life (t1/2) of 29.1 seconds, indicating a rapid exchange of tau molecules within the droplets (Fig. 3B and C). In contrast, the homotypic tau LLPS showed around 79% recovery with a t1/2 of 19.5 s, indicating different dynamics and viscoelastic properties (Fig. S12A and B, ESI†). The decrease in fluorescence recovery and increase in t1/2 suggest rigidification, implying that GA accelerates the maturation of droplets into a gel-like state.29 Further, Young's modulus was assessed by determining the log of Derjaguin, Muller, Toropov (LogDMT) to study rigidification using AFM (Fig. 3D). The homotypic tau LLPS exhibited a logDMT of ∼0.84 Pa, while GA promoted tau LLPS increased to ∼1.36 Pa indicating maturation of droplets (Fig. 3E). DLS study revealed an increased hydrodynamic diameter of GA-promoted tau LLPS, supporting its assembly into larger droplets (Fig. 3F). FTIR spectral changes around 2800–3400 (O–H, N–H stretching), and 1364 cm−1 (–CH3 bending) suggest conformational modulation in tau protein upon LLPS induction (Fig. S13, ESI†). Morphologically, Cryo-TEM characterization revealed tau droplets were promoted in the presence of GA in the solvated state (Fig. 4A). While tau in the presence of AA led to fibril formation, the presence of GA resulted in the coexistence of droplets and short fibrillar structures, indicating GA's capability to modulate AA-induced tau aggregation (Fig. 4B and C). Subsequently, ThT assay revealed GA inhibited AA (185 μM) induced tau (10 μM) aggregation in a dose-dependent manner with 20 μM of GA reducing the fibrillar content to 66% compared to untreated tau (100%) (Fig. 4D). These findings suggest that GA can inhibit fibril formation, despite enhancing the LLPS of tau. GA appears to modulate the intermolecular interactions within tau droplets, leading to changes in the structural dynamics of LLPS, thereby preventing the formation of toxic fibrillar aggregates.29,34 GA was found to be relatively non-toxic to SH-SY5Y neuronal cells up to 100 μM for 24 h (Fig. S14, ESI†). To ascertain the rescue of SH-SY5Y cells from the toxicity induced by tau aggregates, we evaluated the cytotoxicity of AA-induced tau aggregates independently and in the presence of GA. AA-induced tau aggregates reduced cell viability to 79% compared to control (100%). However, GA rescued cells in a dose-dependent manner, with 50 μM of GA increasing the cell viability to 91% (Fig. 4E). Although 10 μM GA promoted LLPS in vitro, no significant rescue was observed, possibly due to interference from proteins in the biological milieu. Higher concentrations promoted stronger condensation, redirecting the toxic pathway. These findings substantiate that GA modulates tau LLPS to drive towards a non-toxic state, highlighting the underexplored therapeutic potential of targeting LLPS in tauopathies. Next, we transfected GFP tau plasmid in SHSY5Y to study the ability of GA to modulate tau LLPS.26 While treatment with congo red (CR) promoted stress induced tau droplets, GA (40 μM) notably decreased the DN indicating their ability to modulate tau LLPS in cells (Fig. 4F and G). Insights gained from our investigations with GA prompted the identification of chemical properties crucial in discerning other biphasic SPs with therapeutic potential. We screened molecules such as DHBA, DP, LD, and SA, possessing phenolic and hydrophobic moieties, similar to GA (Fig. 1A). Akin to GA, biphasic modulation of tau LLPS was observed, validated through turbidity measurements and DIC microscopy for LD and DHBA (Fig. S15–S17, ESI†). These findings affirm the understanding derived from GA studies, facilitating the identification of additional small molecular biphasic modulators of tau LLPS.
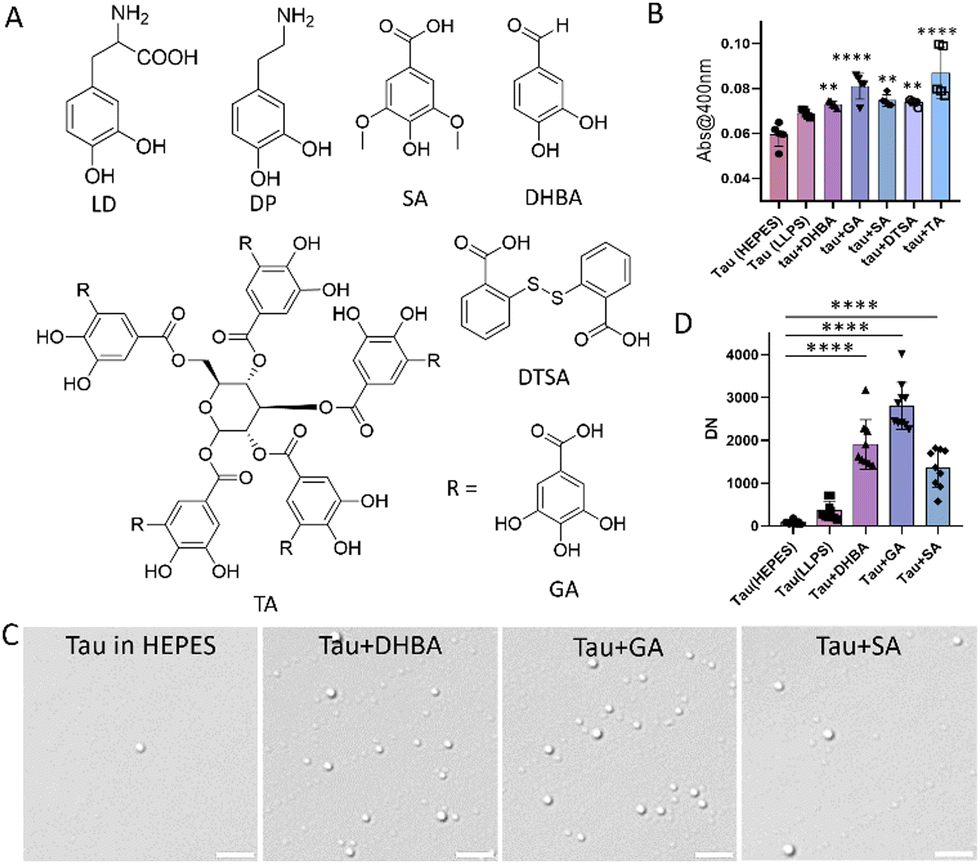 | ||
| Fig. 1 Modulation of tau LLPS by polyphenols. (A) Structures of polyphenols studied, (B) monitoring LLPS induction with turbidity change using SPs. (C) Differential interference contrast (DIC) imaging of tau (5 μM) droplets in the presence of SPs (10 μM) and their quantification represented in (D) (scale bar 5 μm) (DN: droplet number) (statistical analysis conducted using ordinary one-way ANOVA, Tukey's post hoc test *p = 0.05). | ||
 | ||
| Fig. 2 GA Modulates tau LLPS. (A) Turbidity assay of tau with GA, (B) DIC imaging of tau droplets with GA and its quantification represented in (C). (Scale bar 5 μm). (D) Effect on tau LLPS using varying concentrations of GA studied using TEM (scale bar 200 nm). Time-dependent study of GA promoted tau LLPS using (E) DIC quantification and (F) AFM. (Statistical analysis conducted using ordinary one-way ANOVA, Tukey's post hoc test *p = 0.05). | ||
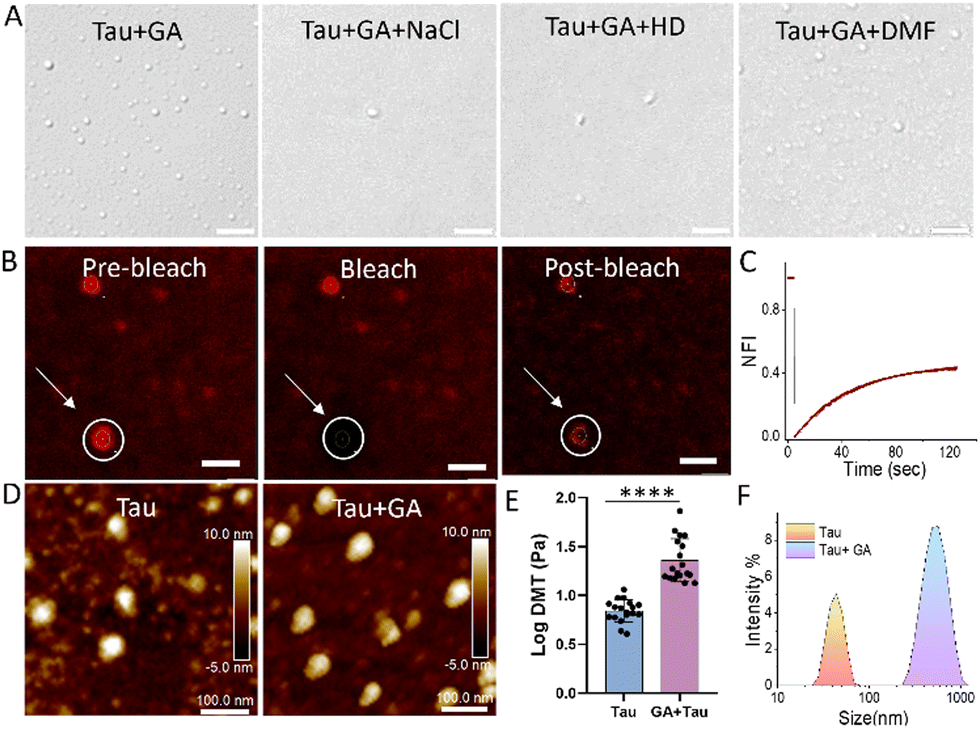 | ||
| Fig. 3 Characterization of GA induced tau LLPS. (A) DIC shows the impact of NaCl, HD, and (DMF) on the modulation of tau LLPS, (scale bar 5 μm). (B) Demonstrates the fluorescence recovery after photobleaching (FRAP) assay of tau LLPS promoted by GA and their (C) subsequent recovery, (scale bar 1 μm). (D) AFM study to monitor morphology and (E) mechanical properties. (F) DLS study indicate tau LLPS promoted by GA (statistics was performed with Student's t-test with ****p < 0.0001). | ||
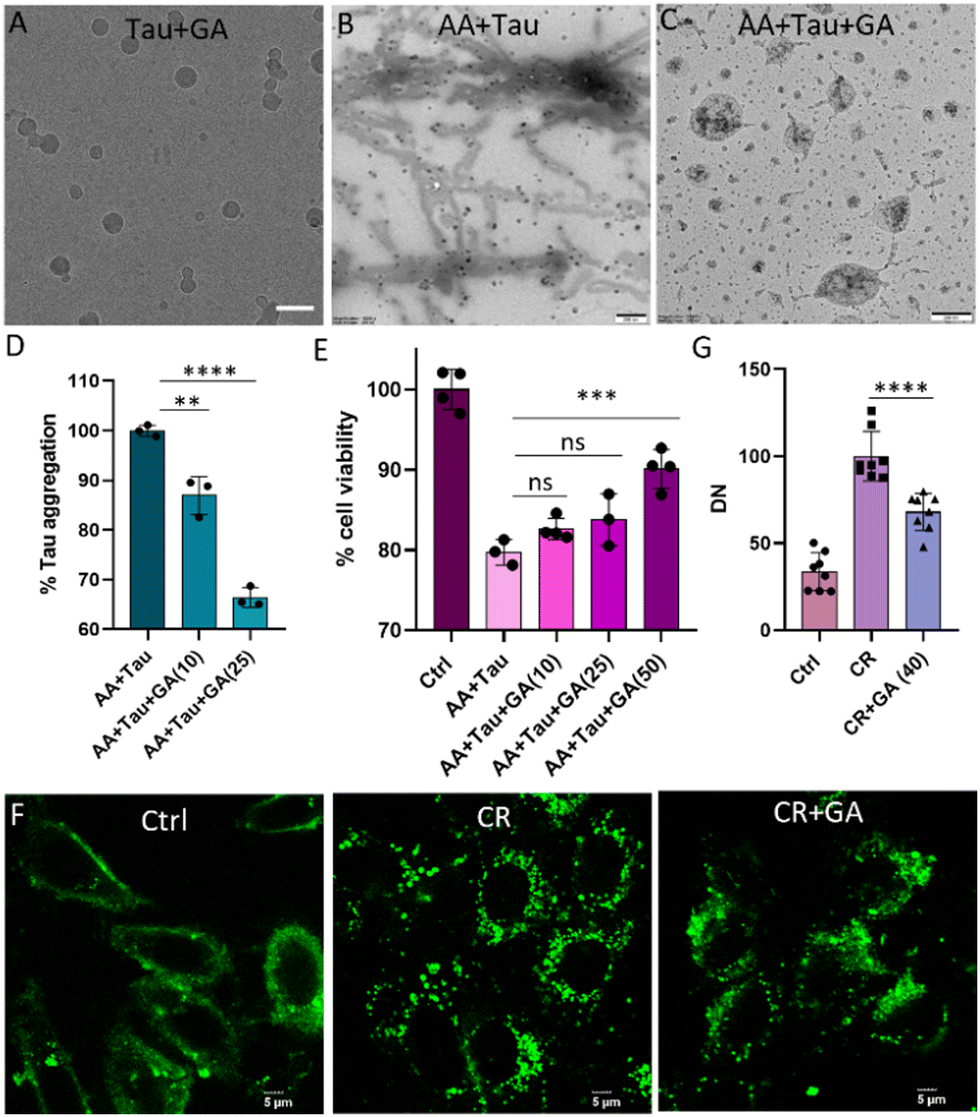 | ||
| Fig. 4 Effect of GA on tau aggregation and toxicity. (A) CryoEM image of tau droplet induced with GA (scale bar 50 nm), TEM image of AA-induced tau aggregation (B) without and (C) with GA (scale bar 200 nm). (D) ThT assay of tau aggregation induced by AA under the influence of GA. (E) MTT assay to monitor the neuronal rescue by GA from tau aggregates toxicity. (F) Confocal image of GFP–Tau droplets and its modulation by GA with its respective quantification in (G) (statistical analysis conducted using ordinary one-way ANOVA, Tukey's post hoc test *p = 0.05). | ||
In conclusion, the investigation of tau LLPS and its regulation is a rapidly evolving field with significant implications for treating neurodegenerative diseases and tauopathies. While efforts have focused on finding compounds to disrupt tau LLPS, emerging evidence highlights the importance of LLPS in normal function. Our study reveals that the natural polyphenol GA acts as a biphasic modulator of tau LLPS, mediated by hydrophobic and electrostatic interactions. This modulation renders GA a transient weak cross-linker, altering tau's metastable state. In contrast to AA-induced toxic fibrillar aggregation, GA promotes the formation of LLPS-driven non-toxic aggregates, protecting neuronal cells from tau toxicity. These findings not only demonstrate the ability of SPs to regulate LLPS but also lay the groundwork for identifying potential leads that could modulate tau LLPS for therapeutic benefits in tauopathies.
We thank JNCASR, core grant (CRG/2020/004594), Science and Engineering Research Board (SERB), New Delhi, India, for the project funding. HM thanks CSIR for fellowship, Shiva Prasad for microscopy experiments.
Conflicts of interest
There are no conflicts to declare.Notes and references
- A. D. C. Alonso, I. Grundke-Iqbal and K. Iqbal, Nat. Med., 1996, 2, 783–787 CrossRef CAS PubMed.
- E.-M. Mandelkow and E. Mandelkow, Trends Cell Biol., 1998, 8, 425–427 CrossRef CAS PubMed.
- T. Govindaraju, Alzheimer's disease: Recent Findings in Pathophysiology, Diagnostic and Therapeutic Modalities, Royal Society of Chemistry, London, 2022 Search PubMed.
- C. Ballatore, V. M.-Y. Lee and J. Q. Trojanowski, Nat. Rev. Neurosci., 2007, 8, 663–672 CrossRef CAS PubMed.
- M. Ramesh and T. Govindaraju, Chem. Sci., 2022, 13, 13657–13689 RSC.
- L. M. Ittner and J. Götz, Nat. Rev. Neurosci., 2011, 12, 67–72 CrossRef PubMed.
- B. Bulic, M. Pickhardt, B. Schmidt, E.-M. Mandelkow, H. Waldmann and E. Mandelkow, Angew. Chem., Int. Ed., 2009, 48, 1740–1752 CrossRef CAS PubMed.
- P. Baruah, H. Moorthy, M. Ramesh, D. Padhi and T. Govindaraju, Chem. Sci., 2023, 14, 9427–9438 RSC.
- J. Chen, W. Ma, J. Yu, X. Wang, H. Qian, P. Li, H. Ye, Y. Han, Z. Su, M. Gao and Y. Huang, J. Agric. Food Chem., 2023, 71, 1982–1993 CrossRef CAS PubMed.
- Q. Zheng, M. T. Kebede, M. M. Kemeh, S. Islam, B. Lee, S. D. Bleck, L. A. Wurfl and N. D. Lazo, Molecules, 2019, 24, 2316–2336 CrossRef CAS PubMed.
- M. Ramesh, A. Acharya, N. A. Murugan, H. Ila and T. Govindaraju, ChemBioChem, 2021, 22, 3348–3357 CrossRef CAS PubMed.
- C. R. Harrington, J. M. D. Storey, S. Clunas and K. A. Harrington, et al. , J. Biol. Chem., 2015, 290, 10862–10875 CrossRef CAS PubMed.
- H. Wood, Nat. Rev. Neurol., 2016, 12, 493 CrossRef CAS PubMed.
- Z. Feng, X. Chen, X. Wu and M. Zhang, J. Biol. Chem., 2019, 294, 14823–14835 CrossRef CAS PubMed.
- B. Wang, L. Zhang, T. Dai, Z. Qin, H. Lu, L. Zhang and F. Zhou, Signal Transduction Targeted Ther., 2021, 6, 290 CrossRef PubMed.
- J. R. Mann and C. J. Donnelly, Neuron, 2021, 109, 2663–2681 CrossRef CAS PubMed.
- S. Ambadipudi, J. Biernat, D. Riedel, E. Mandelkow and M. Zweckstetter, Nat. Commun., 2017, 8, 275 CrossRef PubMed.
- S. Wegmann, B. Eftekharzadeh, K. Tepper and K. M. Zoltowska, et al. , EMBO J., 2018, 37, e98049 CrossRef PubMed.
- Y.-Y. Luo, J.-J. Wu and Y.-M. Li, Chem. Commun., 2021, 57, 13275–13287 RSC.
- H. Ainani, N. Bouchmaa, R. Ben Mrid and R. El Fatimy, Neurobiol. Dis., 2023, 178, 106011 CrossRef CAS PubMed.
- Y. Lin, J. McCarty, J. N. Rauch, K. T. Delaney, K. S. Kosik, G. H. Fredrickson, J.-E. Shea and S. Han, eLife, 2019, 8, e42571 CrossRef PubMed.
- S. Boyko and W. K. Surewicz, Trends Cell Biol., 2022, 32, 611–623 CrossRef CAS PubMed.
- N. M. Kanaan, C. Hamel, T. Grabinski and B. Combs, Nat. Commun., 2020, 11, 2809 CrossRef CAS PubMed.
- J. Wen, L. Hong, G. Krainer, Q.-Q. Yao, T. P. J. Knowles, S. Wu and S. Perrett, J. Am. Chem. Soc., 2021, 143, 13056–13064 CrossRef CAS PubMed.
- W. M. Babinchak, B. K. Dumm, S. Venus, S. Boyko, A. A. Putnam, E. Jankowsky and W. K. Surewicz, Nat. Commun., 2020, 11, 5574 CrossRef CAS PubMed.
- B. Dai, T. Zhong, Z.-X. Chen, W. Chen, N. Zhang, X.-L. Liu, L.-Q. Wang, J. Chen and Y. Liang, J. Biol. Chem., 2021, 297, 101222 CrossRef CAS PubMed.
- P. R. Prince, J. Hochmair, H. Brognaro, S. Gevorgyan and M. Franck, et al. , Sci. Rep., 2023, 13, 3963 CrossRef CAS PubMed.
- M. Ramesh, C. Balachandra, P. Baruah and T. Govindaraju, J. Pept. Sci., 2023, 29, e3465 CrossRef CAS PubMed.
- Y. Huang, J. Wen, L.-M. Ramirez, E. Gümüşdil and P. Pokhrel, et al. , Nat. Commun., 2023, 14, 5444 CrossRef CAS PubMed.
- H. Moorthy, L. P. Datta and T. Govindaraju, Chem. – Asian J., 2021, 16, 423–442 CrossRef CAS PubMed.
- M. B. Avinash and T. Govindaraju, Acc. Chem. Res., 2018, 51, 414–426 CrossRef CAS PubMed.
- C. Yuan, Q. Li, R. Xing, J. Li and X. Yan, Chem, 2023, 9, 2425–2445 CAS.
- R. Chang, C. Yuan, P. Zhou, R. Xing and X. Yan, Acc. Chem. Res., 2024, 57, 289–301 CrossRef CAS PubMed.
- C. Yuan, M. Yang, X. Ren, Q. Zou and X. Yan, Angew. Chem., Int. Ed., 2020, 59, 17456–17460 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Detailed materials, methods, results. See DOI: https://doi.org/10.1039/d4cc00473f |
| This journal is © The Royal Society of Chemistry 2024 |