
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Low Stress Ion Conductance Microscopy of Sub-Cellular Stiffness†
Richard W.
Clarke‡
*a,
Pavel
Novak‡
b,
Alexander
Zhukov
a,
Eleanor J.
Tyler
c,
Marife
Cano-Jaimez
d,
Anna
Drews
a,
Owen
Richards
a,
Kirill
Volynski
d,
Cleo
Bishop
c and
David
Klenerman
*a
aUniversity Chemical Laboratories, Lensfield Road, Cambridge, CB2 1EW, UK. E-mail: rwc25@cam.ac.uk; dk10012@cam.ac.uk
bSchool of Engineering and Materials Science, Queen Mary University of London, Mile End Road, London, E1 4NS, UK
cCentre for Cell Biology and Cutaneous Research, Queen Mary University of London, 4 Newark Street, London, E1 2AT, UK
dUCL Institute of Neurology, Queen Square, London, WC1N 3BG, UK
First published on 17th August 2016
Abstract
Directly examining subcellular mechanics whilst avoiding excessive strain of a live cell requires the precise control of light stress on very small areas, which is fundamentally difficult. Here we use a glass nanopipet out of contact with the plasma membrane to both exert the stress on the cell and also accurately monitor cellular compression. This allows the mapping of cell stiffness at a lateral resolution finer than 100 nm. We calculate the stress a nanopipet exerts on a cell as the sum of the intrinsic pressure between the tip face and the plasma membrane plus its direct pressure on any glycocalyx, both evaluated from the gap size in terms of the ion current decrease. A survey of cell types confirms that an intracellular pressure of approximately 120 Pa begins to detach the plasma membrane from the cytoskeleton and reveals that the first 0.66 ± 0.09 μm of compression of a neuron cell body is much softer than previous methods have been able to detect.
Introduction
In ion conductance microscopy (ICM), insulating surfaces in conducting solution are detected by their slight occlusion of the ion current through the tip aperture of a nanopipet probe.1 A picoampere drop in this nanoampere ion current between the capillary and bath electrodes can be detected within a millisecond using a patch-clamp amplifier, allowing piezoelectric positioning of the nanopipet to map a cell's topography2 and to patch to an exact point of interest.3 It was long thought that ICM imaging exerts almost no stress on a cell simply because the feedback control keeps the tip from making contact with it. However, during approaches to cells the ion current decreases far more slowly with height than its rapid drop next to a hard surface, indicating the glass tip face repels the cell membrane before contact. We were recently able to characterize this interaction by considering the energetic barrier to gigaseal formation in terms of colloid theory4 and now develop the theory to fit data of ion current versus height from deep pushes of cell surfaces. We then show how this understanding allows the quantitative mapping of stiffness across individual cells at low stress, using a variety of cell types in culture – hippocampal neuron (HN)5 cells, a prion protein knockout (Prnp−/−) cell line (HpL),6 and normal, finite lifespan human mammary fibroblast (HMF)7,8 cells.This method is an important technical advance, principally because it allows very soft features of cells to be studied at nanoscale resolution, both in the vertical and lateral directions. To study cells using cantilever techniques, relatively large spheres are typically attached to the tips in order to lower the stress exerted, but this averages out the spatial resolution of differences in stiffness, as well as topography. This same limitation applies to using hydrostatic pressure in ICM as it needs apertures >140 nm9,10 in practice (this reduces blockages from unfiltered particulates carried by the flow), and the flow profile is four times wider than this.9 Fortunately, we find that removing the complication of applying hydrostatic pressure and evaluating the unavoidable forces instead actually makes it more straightforward and less perturbative to image subcellular stiffness, and with higher resolution.
The ability to discern native subcellular structures via stiffness as a second label-free coordinate in addition to topography is itself intrinsically useful, especially given that nanopipets can also deliver reagents to the vicinity11 and make electrochemical measurements.12 The detailed knowledge of the actual structural and mechanical properties in vivo is just as important though, as these determine the overall mechanical properties of the cell, and their rapid and clearly resolved measurement will further the understanding of how cells respond to forces and changes in their environments.
Results & discussion
Approach curves
The tip-face of a typical nanopipet usually remains 50–100 nm distant from the plasma membrane during an ICM scan, not approaching closely enough to make contact with cell-surface macromolecules. To minimise the stress on the cell we avoid the complication of applying hydrostatic pressure9,13 and ensure the slight weight of the column of solution is balanced by surface tension in the capillary.14 Thus in these experiments the stress on most cell types is entirely due to the intrinsic colloidal pressure between the cell surface and the glass tip face. This intrinsic pressure σ varies with the size of the gap between the tip face and the plasma membrane as H/6πg3 where the Hamaker constant H for the glass-cell interaction across physiological saline is estimated to be 4 zJ4 and the gap g is found from an empirical model for the drop in ion current as the nanopipet approaches a hard surface, I = I0(1 − e−xg/r). In such approaches, shown in Fig. 1 and Fig. S1 (ESI†), the aperture radius r is determined from the limiting ion current far from the surface via I0 = πrκV![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) tan(α),15 where the half-cone angle α is 3 degrees, κ is 1.35 S m−1 and V is 200 mV. The tip radius r also determines, along with the empirically determined constant x = 3.6 ± 0.2, the scale of the fall in ion current as the gap narrows. With these values as fixed parameters it is then possible to fit approach data to cells as well, because both ion current and the stresses are fixed functions of the tip-cell gap. For example, the intrinsic stress in terms of the decrease in ion current ΔI is
tan(α),15 where the half-cone angle α is 3 degrees, κ is 1.35 S m−1 and V is 200 mV. The tip radius r also determines, along with the empirically determined constant x = 3.6 ± 0.2, the scale of the fall in ion current as the gap narrows. With these values as fixed parameters it is then possible to fit approach data to cells as well, because both ion current and the stresses are fixed functions of the tip-cell gap. For example, the intrinsic stress in terms of the decrease in ion current ΔI is| σ = H/6π((r/x)ln(I0/ΔI))3 | (1) |
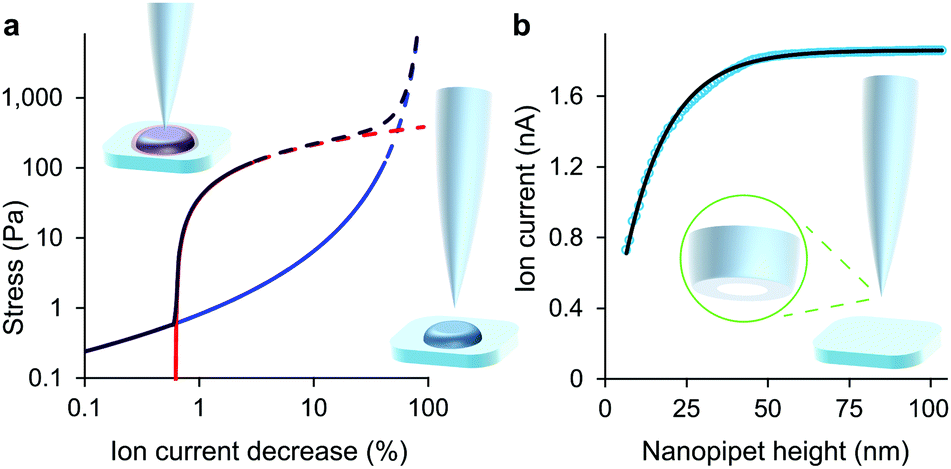 | ||
| Fig. 1 At low decreases in ion current the colloidal interaction between the glass tip face of a nanopipet and the cell membrane exerts a miniscule but quantifiable stress. (a) Stress versus ion current decrease calculated for a 100 nm aperture nanopipet. The total stress (purple) is the sum of the intrinsic stress (blue) and, if present, the direct stress on glycocalyx (red), here set to 70.5 nm; 390 Pa.16 Over 120 Pa (dashes), the cell membrane begins to detach from its anchor-points on the cytoskeleton leading to blebbing.17 Over 8 kPa the tip-face patches to the membrane.4 (b) Ion current through an 84 nm aperture nanopipet approaching a hard flat polystyrene surface. | ||
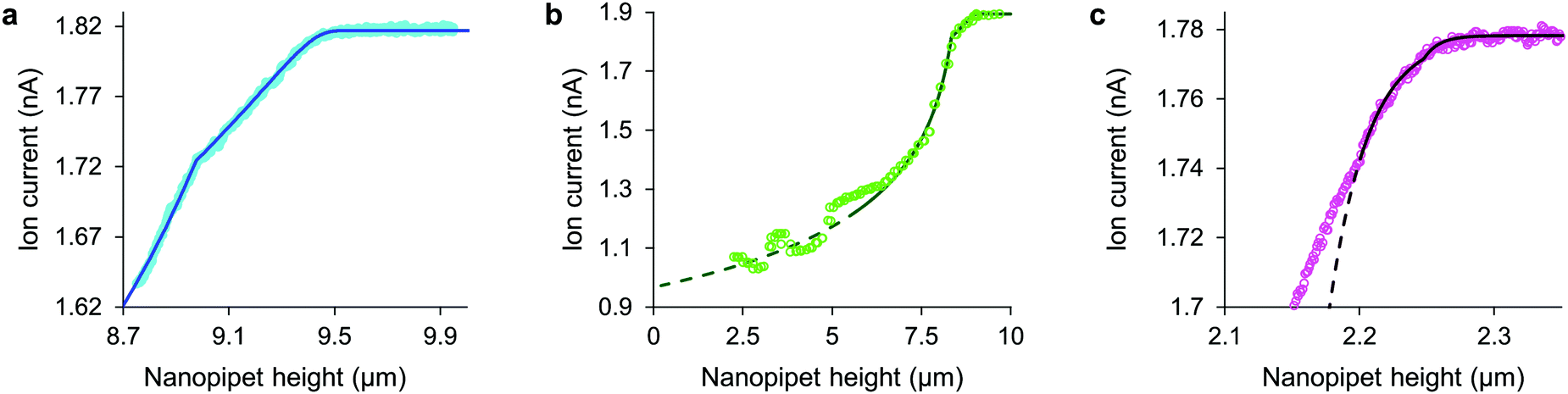 | ||
| Fig. 2 With stress characterized in terms of ion current decrease, approach data fits simple models of cell mechanics: (a) HN cell, apparent stiffness 107 Pa for 0.48 μm, fits 260 Pa, 9.14 μm cortex with 5.5 Pa, 0.29 μm slack. (b) HpL cell, apparent stiffness 52 Pa for 0.70 μm, fits 350 Pa, 8.4 μm cortex with 4 Pa, 0.6 μm slack. (c) HMF cell fits 9.0 kPa, 2.2 μm height with 62 nm glycocalyx. The fit lines are shown dashed at the membrane detachment stress of 120 Pa. | ||
Having detected the plasma membrane at low stress it is remarkable that if our measurements had not pushed far enough to use up the slack there would have been no indication of its existence, for in the initial regime of compression the dual stiffness fit is identical to the uniform model. Its stiffness and rest height parameters for the cortex and slack even combine analytically in the following simple formula to give exactly the same apparent stiffness:
| E = (hC + hS)/((hC/EC) + (hS/ES)) | (2) |
Some other cell types are coated by a porous network of proteoglycans called the glycocalyx. The tip face compresses this elastically when in contact, allowing the stiffness of such cells also to be determined without close approach to the plasma membrane. Any glycocalyx is only strained when the gap g is less than its thickness t, generating a direct stress ς = Y(1 − (g/t)) that adds to the intrinsic pressure on the membrane as shown in Fig. 1. In this paper we take the elastic modulus of HMF glycocalyx Y to be that of human umbilical vein endothelial cell glycocalyx, 390 Pa.16 The decrease in ion current with height is then fully determined by the elastic modulus of the cell and the thickness of the glycocalyx around it. Conversely, these parameters can be inferred in order to fit data of ion current versus tip height, as shown in Fig. 2(c) and Fig. S2 (ESI†). Our HMF approach data fit means of 3.9 ± 0.6 kPa cell stiffness and 70.5 ± 1.6 nm glycocalyx thickness, comparable to the 39.5 nm thickness determined for erythrocytes22 and 3.2–75.0 nm for endothelial cells.23
Although the intrinsic stress increases sharply with ΔI, it fully compresses most cells well before reaching its 8 kPa maximum when, in the absence of glycocalyx, the tip face seals to the membrane.3,4 Thus soft cells like neurons cannot be patched without applying negative hydrostatic pressure, and ion current during approaches is rarely asymptotic to zero. At full compression, where the apical and basal membranes are pushed together against the substrate, sealing would be quickly followed by membrane rupture, at 3 MPa.24 We did not push the cells this far. A much earlier consideration when compressing a cell is the piece-wise detachment of the plasma membrane from its anchor-points on the cytoskeleton, which begins at intracellular pressures of 45–300 Pa.17 This phenomenon allows a cell to accommodate distortion without bursting, and is known as blebbing. Cells actively re-attach folds of plasma membrane to the cytoskeleton, so a live cell is able to wrinkle blebs back into place. However, not exceeding the blebbing stress in the first place maintains a passive elastic response that does not require this energy expenditure. A good estimate of the stress at which these effects typically begin is the log-mean of the above range, 120 Pa. Around this point, where the lines in all figures become dashes, the ion current can decrease slower than expected due to gradual membrane detachment decreasing the effective stiffness, thereby maintaining the tip-cell gap. Any sudden blebbing can reduce the intracellular pressure to such an extent that the gap actually re-widens, whereupon the ion current jumps upwards, as in Fig. 2(b) and Fig. S2 (ESI†). The ion current sometimes decreases faster than expected instead, indicating a second increase in cortical stiffness.
Low stress mapping
In contrast to the approach data which extend to large decreases in ion current, to map cell stiffness we determine the height of each point above the substrate at two low set decreases in ion current, typically only 0.3% and 1.5%. Two fields of nanopipet heights are thus measured at two constant stresses; at a minimal stress of 0.1–10 Pa and at a compressive stress of 1–100 Pa, where both are precisely determined in any particular scan by its specific parameters, described earlier. The nanopipet heights are converted to cell heights by subtracting the tip-cell gap, which typically decreases from 50 to 30 nm for neuronal cells (or from 80 to 50 nm for fibroblasts, where the pipet must be wide enough to detect the cell surface before pushing into the glycocalyx). The stresses and corresponding cell heights are thereby accurately evaluated even though the imaging process only momentarily pushes the cell surface around 100 nm on average at each point. The two simultaneous equations E = Σ1/ε1 = Σ2/ε2, where Σ is the total stress σ + ς and ε denotes strain, then give the cell stiffness as:| E = ((Σ2 − Σ1)h1/(h1 − h2)) + Σ1 |
| = (ΔΣ(z1 − g1)/(Δz − Δg)) + Σ1 | (3) |
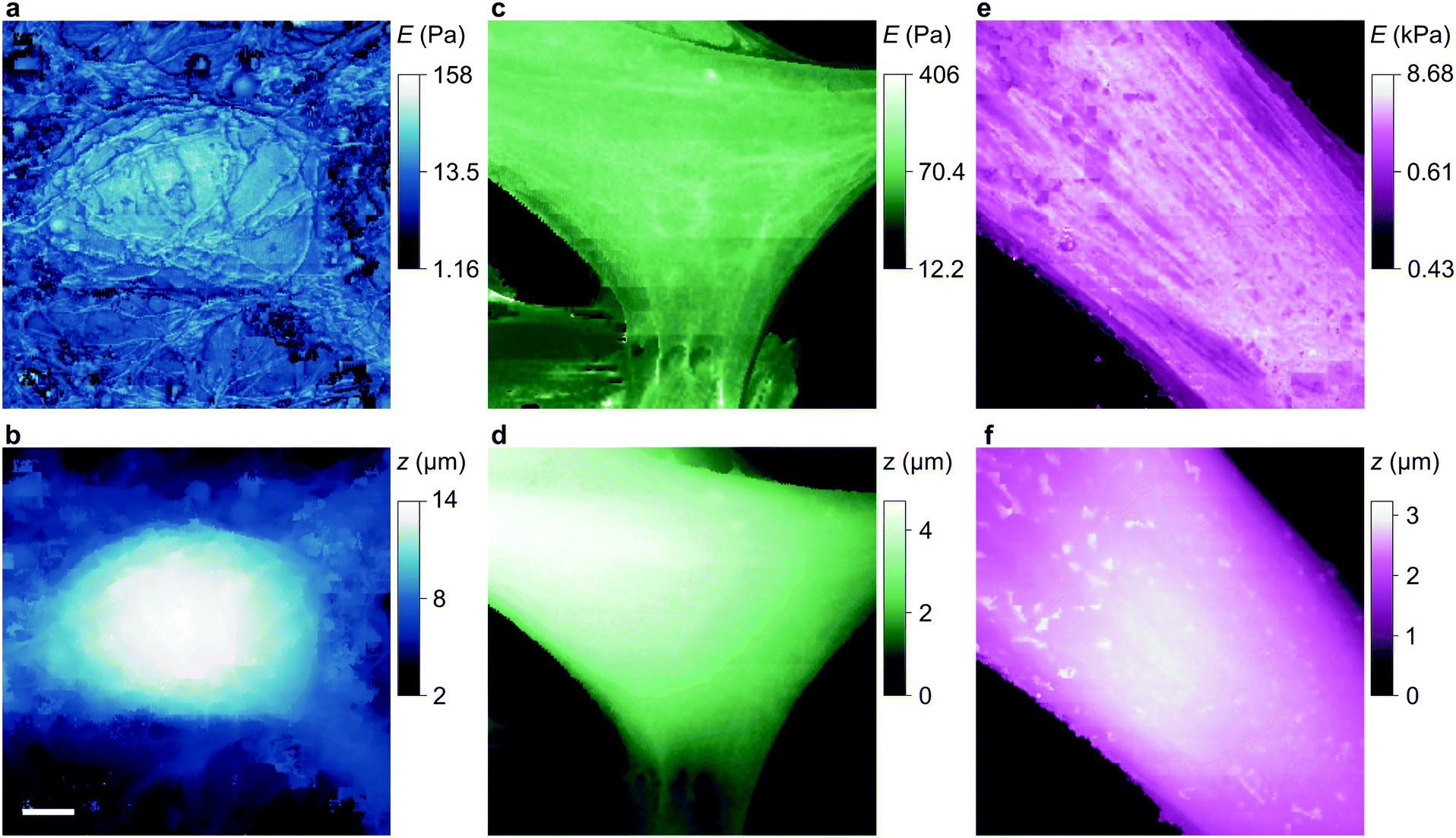 | ||
| Fig. 3 Stiffness and topography of three cell types imaged at high resolution by nanopipet ICM. (a and b) HN neuron mapped by a 110 nm aperture nanopipet at ΔI = 0.3%, 2% exerting stress of 0.29 Pa, 0.96 Pa at tip-cell gaps of 90 nm, 60 nm that typically push the cell soma 0.1–0.2 μm. (c and d) HpL cell mapped by a 52 nm aperture nanopipet at ΔI = 0.6%, 3% exerting stress of 4.27 Pa, 13.26 Pa at gaps of 37 nm, 25 nm. (e and f) HMF fibroblast mapped by a 100 nm aperture nanopipet at ΔI = 0.3%, 3% exerting stress of 0.4 Pa, 122.5 Pa at gaps of 81 nm, 49 nm, resolving the stiffness of stress fibres. The substrate stiffness is masked out in dark blue. The lateral scale bar is 4 μm. | ||
 | ||
| Fig. 4 Stiffness and topography of three cell types imaged at high resolution by nanopipet ICM. (a and b) HN soma mapped by a 62 nm aperture nanopipet at ΔI = 0.5%, 1%, exerting stress of 2.23 Pa, 3.40 Pa at gaps of 46 nm, 40 nm. The lateral scale bar is 1 μm. (c and d) HpL soma mapped by a 52 nm aperture nanopipet at ΔI = 0.6%, 4.2%, exerting stress of 4.27 Pa, 17.95 Pa at gaps of 37 nm, 23 nm. (e and f) Apical area of HMF fibroblast mapped by a 100 nm aperture nanopipet at ΔI = 0.3%, 3%, exerting stress of 0.4 Pa, 122.5 Pa at gaps of 81 nm, 49 nm. Cell surface structures, probably endocytotic events, are visible in the stiffness map but not in the topography. | ||
Note that the stiffness maps of fibroblasts need an independent estimate of glycocalyx stiffness and also its thickness as determined from approach data. Glycocalyx stiffness does depends on cell type; it is 250 Pa for pulmonary endothelial cells for example,25 but the dominant direct stress is linear in this parameter so changing it does not affect the contrast in stiffness that will usually be of primary interest.
To compare our measurements with other techniques that have assayed cell body stiffness we calculated the mean stiffness of somatal regions in our scans as identified by topography, demonstrated in Fig. S3 (ESI†). The hippocampal neurons have an apparent cell body stiffness of 56 ± 9 Pa, corresponding to a cortical stiffness of 310 ± 109 Pa. When assessed by 6 μm diameter polystyrene spheres on a cantilever tip the stiffness of similar cell bodies was reported to be 900 Pa,26 suggesting the cantilever spring constant was too high to detect the initial slack and instead measured the cortical stiffness directly. Some HN cells we measured did have a cortex this stiff but others were much softer; we suspect that in earlier cantilever studies this softer subpopulation would have been flattened against the substrate and missed. HpL cells had an apparent cell body stiffness of 64 ± 4 Pa, corresponding to a cortical stiffness of 702 ± 22 Pa. The stiffness we find for HMF cells, 2.25 ± 0.27 kPa, is comparable to an average stiffness from force microscopy of fibroblasts,27,28 2.89 ± 0.28 kPa. These cells are stiffened by the enhanced lateral force transmission of the numerous stress fibres29 seen in Fig. 3(e) and 4(e).
As a reference for future studies, the dependencies and limiting factors of the stresses in ICM are illustrated in Fig. S8 (ESI†). Each curve begins at the minimum detectable ion current decrease in 1 ms, calculated for a signal to noise ratio of three times the thermal noise, 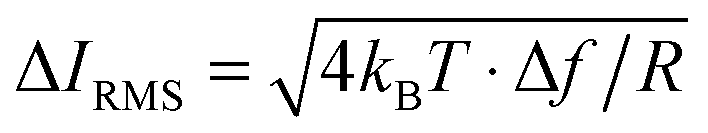 .30 These values increase for smaller aperture diameters but it would be possible to detect smaller changes in ion current for higher resistance nanopipets by extending the acquisition time. The bandwidth Δf = 1 kHz corresponds to the rate of data acquisition typically necessary for imaging experiments.
.30 These values increase for smaller aperture diameters but it would be possible to detect smaller changes in ion current for higher resistance nanopipets by extending the acquisition time. The bandwidth Δf = 1 kHz corresponds to the rate of data acquisition typically necessary for imaging experiments.
Conclusions
These are general methods for assaying and imaging cell stiffness but they have already identified here some specific features of interest. We have shown how to determine the thickness of the glycocalyx from approach data, and have found that some subcellular structures exhibit strong contrast in stiffness but none in topography. We have also identified that the initial deformation of most points of the plasma membrane of neurons is extremely soft, indicating that we must often be encountering the spaces between its non-tethered points and the cortical cytoskeleton, and/or conformational slack in the cytoskeleton itself.Overall, these results demonstrate that it is possible to map the stiffness of cells at very high resolution, both laterally and vertically, without the considerable effort of modifying ICM apparatus to apply hydrostatic pressure. The absence of flow also allows narrower nanopipets to be used that would otherwise be prone to blockages, and for which the forces we describe would have to be evaluated in any case. Further advantages stem from minimizing the offset from the tip-face to the cell surface – if applying hydrostatic pressure this offset is necessarily larger to accommodate the flow profile, which lowers resolution and begins pushing the cell before its surface is detected. The equations developed here also indicate that it may be possible to patch hard cells without the requirement of applying negative hydrostatic pressure.
Besides its ability to map a vast range of stiffness at the nanoscale, stress-quantitative ICM will now enable many other interesting studies of live cells, including fundamentally non-invasive assays of differentiation, subcellular response, and mechanosensation. It will be possible for example to assay the exact stresses at which mechanosensitive ion channels open. Thus this advance in understanding of ICM greatly increases its versatility for nanoscale biophysics and the study of cellular mechanics.
Author contributions
R. W. C. developed the models and experimental design; P. N. and R. W. C. designed the apparatus and scanned the cells; P. N., R. W. C. and A. Z. coded the controllers; P. N., A. Z. and O. R. recorded the approaches; R. W. C., P. N. and A. Z. processed the data; M. C.-J. and K. V. contributed the HN cells; A. D. cultured the HpL cells; E. J. T. and C. B. contributed the HMF cultures; and R. W. C. and D. K. directed the research.Acknowledgements
We thank Prof. P. St George-Hyslop and Prof. Dr A. Aguzzi for the HpL cells, Dr M. Stampfer for the HMF cells, and Dr S. Antolin for comments. This work was supported by the BBSRC (BB/L006227/1), EPSRC (EP/H01098X/1) and MRC (G0701057, MR/K501372/1). A. D. was funded by a Herchel Smith Postdoctoral Fellowship and O. R. by the RSC Analytical Chemistry Trust Fund.References
- P. Hansma, B. Drake, O. Marti, S. Gould and C. Prater, Science, 1989, 243, 641–643 CAS.
- P. Novak, C. Li, A. I. Shevchuk, R. Stepanyan, M. Caldwell, S. Hughes, T. G. Smart, J. Gorelik, V. P. Ostanin and M. J. Lab, Nat. Methods, 2009, 6, 279–281 CrossRef CAS PubMed.
- P. Novak, J. Gorelik, U. Vivekananda, A. I. Shevchuk, Y. S. Ermolyuk, R. J. Bailey, A. J. Bushby, G. W. Moss, D. A. Rusakov and D. Klenerman, Neuron, 2013, 79, 1067–1077 CrossRef CAS PubMed.
- R. W. Clarke, A. Zhukov, O. Richards, N. Johnson, V. Ostanin and D. Klenerman, J. Am. Chem. Soc., 2012, 135, 322–329 CrossRef PubMed.
- G. A. Banker and W. M. Cowan, Brain Res., 1977, 126, 397–425 CrossRef CAS PubMed.
- C. Kuwahara, A. M. Takeuchi, T. Nishimura, K. Haraguchi, A. Kubosaki, Y. Matsumoto, K. Saeki, Y. Matsumoto, T. Yokoyama and S. Itohara, Nature, 1999, 400, 225–226 CrossRef CAS PubMed.
- S. R. Romanov, B. K. Kozakiewicz, C. R. Holst, M. R. Stampfer, L. M. Haupt and T. D. Tlsty, Nature, 2001, 409, 633–637 CrossRef CAS PubMed.
- M. G. Overhoff, J. C. Garbe, J. Koh, M. R. Stampfer, D. H. Beach and C. L. Bishop, Nucleic Acids Res., 2014, 42, 1606–1618 CrossRef CAS PubMed.
- J. Rheinlaender and T. E. Schäffer, Soft Matter, 2013, 9, 3230–3236 RSC.
- J. Rheinlaender, S. Vogel, J. Seifert, M. Schaechtele, O. Borst, F. Lang, M. Gawaz and T. E. Schaeffer, Thromb. Haemostasis, 2015, 113, 305–311 CrossRef PubMed.
- B. Babakinejad, P. Jönsson, A. López Córdoba, P. Actis, P. Novak, Y. Takahashi, A. Shevchuk, U. Anand, P. Anand and A. Drews, Anal. Chem., 2013, 85, 9333–9342 CrossRef CAS PubMed.
- Y. Takahashi, A. I. Shevchuk, P. Novak, Y. Murakami, H. Shiku, Y. E. Korchev and T. Matsue, J. Am. Chem. Soc., 2010, 132, 10118–10126 CrossRef CAS PubMed.
- D. Sánchez, N. Johnson, C. Li, P. Novak, J. Rheinlaender, Y. Zhang, U. Anand, P. Anand, J. Gorelik and G. I. Frolenkov, Biophys. J., 2008, 95, 3017–3027 CrossRef PubMed.
- M. Pellegrino, P. Orsini, M. Pellegrini, P. Baschieri, F. Dinelli, D. Petracchi, E. Tognoni and C. Ascoli, Neurosci. Res., 2011, 69, 234–240 CrossRef PubMed.
- T. Chowdhury, J. Phys. E: Sci. Instrum., 1969, 2, 1087 CAS.
- K. Bai and W. Wang, J. R. Soc., Interface, 2012, 9, 2290–2298 CrossRef CAS PubMed.
- G. T. Charras, M. Coughlin, T. J. Mitchison and L. Mahadevan, Biophys. J., 2008, 94, 1836–1853 CrossRef CAS PubMed.
- A. Sakudo and T. Onodera, Front. Cell Dev. Biol., 2015, 2, 75 Search PubMed.
- L. Xie, H. Kang, Q. Xu, M. J. Chen, Y. Liao, M. Thiyagarajan, J. O'Donnell, D. J. Christensen, C. Nicholson and J. J. Iliff, Science, 2013, 342, 373–377 CrossRef CAS PubMed.
- S. Dreha-Kulaczewski, A. A. Joseph, K.-D. Merboldt, H.-C. Ludwig, J. Gärtner and J. Frahm, J. Neurosci., 2015, 35, 2485–2491 CrossRef CAS PubMed.
- J. J. Iliff, M. Wang, D. M. Zeppenfeld, A. Venkataraman, B. A. Plog, Y. Liao, R. Deane and M. Nedergaard, J. Neurosci., 2013, 33, 18190–18199 CrossRef CAS PubMed.
- G. Geyer and J. Makovitzky, J. Microsc., 1980, 119, 407–414 CrossRef CAS PubMed.
- P. Janczyk, S. Hansen, M. Bahramsoltani and J. Plendl, J. Electron Microsc., 2010, 59, 291–298 CrossRef CAS PubMed.
- D. Needham and R. S. Nunn, Biophys. J., 1990, 58, 997 CrossRef CAS PubMed.
- R. O'Callaghan, K. M. Job, R. O. Dull and V. Hlady, Am. J. Physiol.: Lung Cell. Mol. Physiol., 2011, 301, L353–L360 CrossRef PubMed.
- Y.-B. Lu, K. Franze, G. Seifert, C. Steinhäuser, F. Kirchhoff, H. Wolburg, J. Guck, P. Janmey, E.-Q. Wei, J. Käs and A. Reichenbach, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 17759–17764 CrossRef CAS PubMed.
- M. Plodinec, M. Loparic, C. A. Monnier, E. C. Obermann, R. Zanetti-Dallenbach, P. Oertle, J. T. Hyotyla, U. Aebi, M. Bentires-Alj and R. Y. Lim, Nat. Nanotechnol., 2012, 7, 757–765 CrossRef CAS PubMed.
- R. Mahaffy, C. Shih, F. MacKintosh and J. Käs, Phys. Rev. Lett., 2000, 85, 880 CrossRef CAS PubMed.
- C. P. Brangwynne, F. C. MacKintosh, S. Kumar, N. A. Geisse, J. Talbot, L. Mahadevan, K. K. Parker, D. E. Ingber and D. A. Weitz, J. Cell Biol., 2006, 173, 733–741 CrossRef CAS PubMed.
- H. Nyquist, Phys. Rev., 1928, 32, 110–113 CrossRef CAS.
Footnotes |
| † Electronic supplementary information (ESI) available: Supplementary methods and expanded data presentations of approach curves, stiffness maps, and nanopipet stresses and forces. See DOI: 10.1039/c6sm01106c |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2016 |