
Bioimaging of the elemental distribution in cocoa beans by means of LA-ICP-TQMS
 ac
Uwe
Karst
*a
ac
Uwe
Karst
*a
Abstract
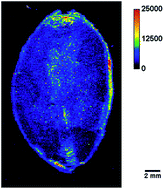
Cocoa beans (Theobroma cacao L.) are a key raw material for the confectionary industry. Therefore, their ingredients, nutrients as well as contaminants, have to be thoroughly analysed. In this work, a method for spatially resolved analysis of the tissue specific nutrient and contaminant distribution in cocoa beans by laser ablation-inductively coupled plasma-triple quadrupole mass spectrometry (LA-ICP-TQMS) is presented. This method provides high spatial resolution for fine structures and a wide linear range to detect macronutrients and lower concentrated toxic elements – such as Cd – in a single experiment. By using a triple quadrupole analyser, elements that are subject to isobaric or polyatomic interferences in ICP-MS analysis, such as Ca or Fe, can also be determined accurately. The distribution of Na, Mg, P, K, Ca, Fe, Cu, Zn, Cd and Pb in cocoa beans was successfully visualized for the first time. The method represents a new approach for studying contaminants in cocoa beans and ultimately the development of innovative strategies to minimize contamination in the product.
- This article is part of the themed collection: Atomic spectrometry for the analysis of biological samples
 Please wait while we load your content...
Please wait while we load your content...

