
Bisphenol analogues differently affect human islet polypeptide amyloid formation†
Lizi Huanga,
Mingyan Liaoa,
Xin Yanga,
Hao Gonga,
Liang Maa,
Yudan Zhaoa and
Kun Huang*ab
aTongji School of Pharmacy, Huazhong University of Science and Technology, Wuhan, China 430030. E-mail: kunhuang2008@hotmail.com
bCenter for Biomedicine Research, Wuhan Institute of Biotechnology, Wuhan, China 430075
First published on 12th January 2016
Abstract
Bisphenols (BPs) are a group of chemicals with two hydroxyphenyl functionalities. BPs analogues including bisphenol A (BPA), bisphenol B (BPB), bisphenol F (BPF), bisphenol S (BPS), bisphenol AF (BPAF), bisphenol AP (BPAP), tetrabromobisphenol A (TBBPA) and tetrachlorobisphenol A (TCBPA) are widely used in the production of plastic materials. There has been a growing concern worldwide about environmental BPs exposure due to their potential adverse effects on human health. Deposition of β-cell toxic misfolded human islet amyloid polypeptide (hIAPP), a hormone biosynthesized and secreted by the pancreatic β-cells, is a cardinal finding in type 2 diabetes mellitus (T2DM). Epidemiological studies suggest BPA exposure is positively associated with the incidence of T2DM, which may be due to the exacerbated toxic aggregation of hIAPP induced by BPA as we previously reported. Here, we demonstrate that bisphenol analogues showed different effects on hIAPP amyloid formation. Unlike the accelerating effects of BPA, four BPs analogues (BPAF, BPAP, TBBPA and TCBPA) showed inhibitory effects on hIAPP aggregation and decreased the membrane disruption capacity of hIAPP. Our results also explained a possible relationship between the spatial structure and the inhibitory capacities of BPs analogues.
1. Introduction
Proteins transform from native conformations into highly ordered fibrillar aggregates under certain circumstances, which may lead to pathological conditions ranging from neurodegenerative disorders to systemic amyloidoses, including Alzheimer's disease (AD),1,2 Parkinson's disease (PD)3 and type 2 diabetes mellitus (T2DM).4,5Human islet amyloid polypeptide (hIAPP) is a 37 residue hormone that is synthesized in pancreatic β-cells and co-secreted with insulin (Fig. 1A).6 Despite its important physiological functions, hIAPP has a high intrinsic propensity to misfold into toxic oligomers and linear fibrils.7 The depletion of islet β-cells correlates strongly with the formation of hIAPP amyloid fibrils.6 Thus, preventing the formation of toxic hIAPP amyloid has been viewed as a plausible therapeutic approach for T2DM.8,9
 | ||
| Fig. 1 Structure of hIAPP and BPA and its analogues. (A) Primary sequence of hIAPP with a disulfide bridge between Cys-2 and Cys-7. (B) Chemical structures of BPA and its analogues. | ||
Bisphenol A (BPA, 2,2-bis-(4-hydroxydiphenyl)-propane) has been widely used in the production of polycarbonate plastics and epoxy resins for decades (Fig. 1B).10 BPA exposure causes many health problems even at very low doses, including breast and prostate cancer, metabolic syndrome, obesity, and subfertility.11 Therefore, the European Food Safety Authority, US Environmental Protection Agency and Health Canada have placed restrictions on BPA application. Nowadays, novel bisphenol analogues, such as bisphenol S (BPS, 4,4′-sulfonyldiphenol), bisphenol F (BPF, 4,4′-methylenebisphenol), bisphenol B (BPB, 2,2-bis-(4-hydroxyphenyl)-butane), bisphenol AF (BPAF or hexafluorobisphenol A) and bisphenol AP (BPAP, 4,4′-(1-phenylethylidene)-bisphenol) have been developed for different purposes (Fig. 1B). For example, BPS is used as an alternative to BPA in the production of thermal paper,12 BPB, BPF, BPAF and BPAP are used in the production of polycarbonate plastic and resins.13 Tetrachlorobisphenol A (TCBPA) and tetrabromobisphenol A (TBBPA) are widely used as reactive flame retardants in resins and polycarbonate plastics (Fig. 1B).14 We previously reported that BPA could accelerate the toxic amyloid formation of hIAPP, which may eventually contribute to the onset of T2DM.15 In this study, we compared eight BPA analogues on their effects on the toxic amyloid formation, and discussed the possible mechanism behind their different actions.
2. Results
2.1. BPA accelerated the amyloid formation of hIAPP, while BPAF, BPAP, TBBPA, TCBPA strongly inhibited this process
Thioflavin-T (ThT) is a fluorescence dye binds specifically to β-sheet structures and is widely used in probing the emerging β-sheet structures during amyloid formation.16 hIAPP was incubated with BPs analogues at molar ratio of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 or 1:10 at 25 °C for 2 h with continuously scanning of the emission fluorescence, respectively. The emission of hIAPP alone reached a steady state after about 1 h of incubation. The ThT fluorescence assay suggests that BPA showed no effect on β-structure formation at a 1:1 ratio, but had a strong acceleration effect at 10:1, which is consistent with our previous report (Fig. 2A).15 In contrast, BPF and BPS showed no effect on amyloid formation even at molar ratio of 10:1 (Fig. S1B & S1C†), BPB partially inhibited the aggregation at a high molar ratio of 10:1, whereas BPAF, BPAP, TBBPA and TCBPA showed inhibitory effects (Fig. 2C, S1A & S1D–F†). At a 10:1 molar ratio, the presence of BPB and BPAP decreased the maximum ThT intensity of hIAPP by 40% to 50% compared to the control (Fig. S1A & S1E†), whereas BPAF, TBBPA and TCBPA almost completely inhibited amyloid formation of hIAPP (Fig. 2B and C, S1D & S1F†).
:1 or 1:10 at 25 °C for 2 h with continuously scanning of the emission fluorescence, respectively. The emission of hIAPP alone reached a steady state after about 1 h of incubation. The ThT fluorescence assay suggests that BPA showed no effect on β-structure formation at a 1:1 ratio, but had a strong acceleration effect at 10:1, which is consistent with our previous report (Fig. 2A).15 In contrast, BPF and BPS showed no effect on amyloid formation even at molar ratio of 10:1 (Fig. S1B & S1C†), BPB partially inhibited the aggregation at a high molar ratio of 10:1, whereas BPAF, BPAP, TBBPA and TCBPA showed inhibitory effects (Fig. 2C, S1A & S1D–F†). At a 10:1 molar ratio, the presence of BPB and BPAP decreased the maximum ThT intensity of hIAPP by 40% to 50% compared to the control (Fig. S1A & S1E†), whereas BPAF, TBBPA and TCBPA almost completely inhibited amyloid formation of hIAPP (Fig. 2B and C, S1D & S1F†).
 | ||
| Fig. 2 Effects of BPA analogues on hIAPP aggregation. ThT-fluoresence of hIAPP with different molar ratio of compounds, (A) BPA; (B) TBBPA; (C) the relative fluorescence intensity of hIAPP with 10 molar ratio of compounds at 2 h. Concentration of hIAPP was 10 μM. *P < 0.05; **P < 0.01; ***P < 0.001. | ||
The morphology of hIAPP aggregates in the presence and absence of BPs analogues was observed by TEM as we previously reported.17 After 1 h of incubation, hIAPP alone showed protofibrils and typical extensive linear mature fibrils, respectively (Fig. S2A†). In the presence of BPA, the linear mature fibrils appeared at 1 h (Fig. S2B†). BPAF attenuated oligomer and fibril formation, no protofibril was observed at 1 h (Fig. S2F†), short fibrils were detected at 2 h (Fig. 3F). BPB, BPF, BPS showed no effect on the formation of hIAPP amyloid compared to the control group (Fig. S2C–E,† and 3C–E). In the presence of BPAP, TBBPA and TCBPA, the amyloid formation seemed to be completely inhibited as no obvious aggregate was observed (Fig. 3G–I, S2G–I†). These observations are consistent with the results obtained from the ThT fluorescence assays.
 | ||
| Fig. 3 TEM images of hIAPP co-incubated with different compounds at 2 h. (A) hIAPP; (B) BPA; (C) BPB; (D) BPF; (E) BPS; (F) BPAF; (G) BPAP; (H) TBBPA; (I) TCBPA. Concentration of hIAPP and compounds was 10 μM and 100 μM, respectively. Scale bar represents 500 nm. | ||
2.2. Except for BPA and BPB, the other six analogues delayed the secondary structure transition of hIAPP
To gain insight into the effects of BPs analogues on the secondary structures of hIAPP aggregation, far-UV CD spectroscopy was applied to monitor conformational changes over time. The CD spectrum of hIAPP was characteristic of predominant random coil structure at the beginning of incubation (Fig. 4A), which was consistent with a previous report.18 After 20 min of incubation, an intensity increase was observed for a negative band between 210–220 nm, which represents a conversion from random coil to β-sheet-rich structure (Fig. 4A). A single negative peak near 220 nm was observed after 40 min incubation as a typical sign of β-structures (Fig. 4A). | ||
| Fig. 4 Far-UV CD spectra of hIAPP with different compounds. Far-UV circular dichroism spectra of sample groups (A–F), and percentages of secondary structure contents at 0 min, 20 min, 60 min as calculated by the program CDPro (G). Concentration of hIAPP and compounds was 20 μM and 200 μM, respectively. | ||
When BPs were added to hIAPP at a molar ratio of 10:1, all tested BPs analogues, except for BPA and BPB, showed different degrees of inhibition effects. Among them, TBBPA and TCBPA were the strongest. BPA accelerated the secondary structure transition of hIAPP (Fig. 4B) while BPB showed no effect on this process (Fig. S3A†). In the presence of BPF, BPS, BPAF and BPAP, the negative band between 210–220 nm appeared at 40 min, and the negative peak near 220 nm appeared at 60 min, which were 20 minutes later compared to the spectra of hIAPP alone (Fig. S3B & S3C,† 4C and D). Furthermore, TBBPA and TCBPA had a strong inhibition capacity on the secondary structure transition of hIAPP (Fig. 4G) since the CD spectra were remained at the original state during 90 min incubation when a 10:1 ratio of these analogues were added to hIAPP (Fig. 4E–F), which was consistent with the quantitative analysis with the program CDPro (Fig. 4G).
2.3. BPAF, BPAP, TBBPA, TCBPA inhibited both oligomer and fibril formation
OC and A11 are polyclonal antibodies that recognize generic epitopes or a peptide backbone epitope that are common to amyloid fibrils or oligomers.19,20 Dot-blot assays probed with the A11 or OC antibody showed that during 2 h incubation, the intensities of oligomer and fibril spots of the hIAPP co-incubated with BPA were stronger than the control, suggesting BPA accelerated hIAPP aggregation (Fig. 5A and B), whereas BPB, BPF and BPS showed no obvious effect (Fig. 5A). In contrast, the presence of a 10:1 ratio of BPAF, BPAP, TBBPA and TCBPA significantly decreased the formation of both oligomers and fibrils compared to the control group (Fig. 5A and B).
 | ||
| Fig. 5 Effects of BPA analogues on hIAPP oligomerization and fibrillation detected by dot blot. (A) 10 μM of hIAPP monomer was incubated at 25 °C in the absence (hIAPP only) or presence of 10× BPA and its analogues (BPB, BPF, BPS, BPAF, BPAP, TBBPA and TCBPA) for up to 2 h. For each antibody, all samples were spotted onto one nitrocellulose membrane. Each membrane was immuno-stained with the OC or A11 antibody. (B) Quantification of the effect of bisphenols on hIAPP aggregation. Due to different film exposure times used for each individual test, no standard deviation can be obtained. | ||
The aggregation process of hIAPP was further monitored by dynamic light scattering (DLS). The average diameter of hIAPP was about 469 nm and the hydrodynamic diameter distribution was mainly at 204 and 1010 nm at 0 min (Fig. 6A). After 15 min incubation, the average diameter of hIAPP alone was increased from 469 nm to 4344 nm with a large portion of particles distribute at 5887 nm (Fig. 6A). In the presence of BPA, particles with the size above 10000 nm was detected (which is the detection limit of the equipment, data not shown) during the same incubation time. In comparison, 10-fold of BPA significantly accelerated the aggregation (Fig. 6B). BPF and BPS showed no inhibitory effect, while BPAF, BPAP, TBBPA, TCBPA all showed remarkable inhibitory effects at a molar ratio of 10:1, with the average particle size remained under 2000 nm at 15 min (Fig. 6F–I). For BPB, a weak inhibition effect was observed (Fig. 6B), which agrees with the results of ThT and TEM.
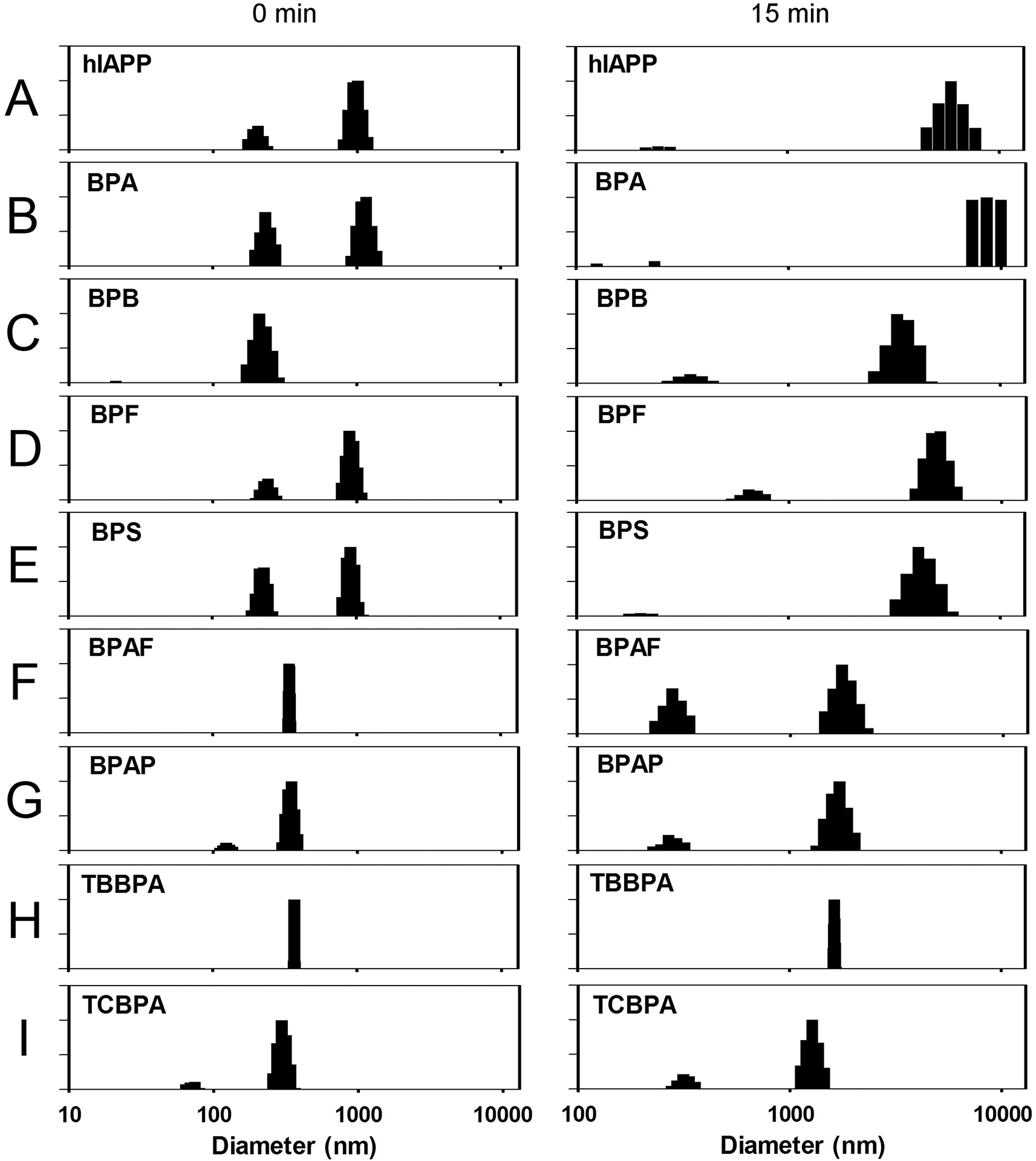 | ||
| Fig. 6 Dynamic light scattering detected hIAPP particle distribution. (A) 10 μM hIAPP. hIAPP with 10-fold (B) BPA; (C) BPB; (D) BPF; (E) BPS; (F) BPAF; (G) BPAP; (H) TBBPA; (I) TCBPA. After 15 min, the average particle size of hIAPP co-incubated with BPA reached 10000 + nm, which was beyond the measuring range of the instrument. | ||
Since many organic chemicals can self-aggregate into colloidal aggregates in aqueous solution and further affect amyloid formation.21 To evaluate this possibility, we used the Aggregator Advisor program (http://advisor.bkslab.org/) to predict whether these bisphenol analogues have the propensity to self-aggregate.21 The prediction suggested six analogues (BPA, BPB, BPAF, BPAP, TBBPA and TCBPA) have calculated logP values > 3, and three of them (BPA, BPAP, TBBPA) are similar to previously reported aggregators by the Aggregator Advisor (Table S1†), indicating their potency to self-aggregate. We then used DLS to continuously detect any self-aggregates of these bisphenol analogues for 24 h, but no aggregates were detected (Table S1†).
2.4. BPB, BPAF, BPAP, TBBPA and TCBPA decreased the membrane disruption ability of hIAPP
It has been reported that hIAPP oligomers and mature fibrils disrupt β-cell membranes and release intracellular contents, which results in β-cell apoptosis and eventually diabetes.22,23 The dye leakage assays were performed by using artificial micelles containing fluorescence dye to probe the membrane disruption capacity of hIAPP. POPG vesicles treated with 0.2% (v/v) Triton-X 100 (complete membrane leakage, set as 100% disruption) were measured as the positive control. Control studies suggested BPs analogues alone only caused mild membrane disruption at 10 μM concentration (Fig. 7A). The addition of 1 μM hIAPP disrupted the membrane by 31.5 ± 0.6% (P < 0.0001, Fig. 7A). In the presence of 10 μM BPA, the membrane disruption was increased to 35.6 ± 0.6% (P < 0.0001). At 10 μM concentration, BPF and BPS showed no effect on membrane disruption during the period of incubation, while the presence of BPB, BPAF, BPAP, TBBPA and TCBPA decreased the membrane disruption to 22.0 ± 0.8%, 11.9 ± 0.9%, 12.3 ± 0.9%, 10.0 ± 0.9% and 11.5 ± 0.9%, respectively (all P < 0.0001, Fig. 7A). The effects of BPA analogues on hIAPP aggregation induced cytotoxicity were further studied on BxPC3 cells, a human pancreatic tumor cell line, by MTT assays. Surprisingly, the presence of bisphenol analogues showed no obvious toxic effect on cell viabilities, with some of these analogues even showed weak proliferation of the cells; moreover, all these analogues showed no significant protective effect on cell viabilities (Fig. S4†), possibly due to the reported estrogenic effects of BPA analogues.24–27 Furthermore, the haemolysis analysis also suggested these BP analogues showed complex effects on haemolysis, even for those analogues showed inhibitory effects on hIAPP amyloid formation in vitro (Fig. S5†), suggesting these analogues cannot be simply regarded as bio-safe. | ||
| Fig. 7 Dye leakage effects of hIAPP co-incubated with BPA and its analogues. (A) Fluorescence dye leakage levels of hIAPP in the presence of different compounds. 0.2% Triton X-100 treated carboxyfluorescein-containing POPG micelles were used as the positive control. (B) BPA and its analogues treated carboxyfluorescein-containing POPG micelles were used as the negative controls. *P < 0.05; **P < 0.01; ***P < 0.001. | ||
3. Discussion
The gradual loss of pancreatic β-cells caused by hIAPP amyloid deposits in the islets of Langerhans has been thought as an importance cause of type 2 diabetes mellitus.28,29 Therefore, inhibition of hIAPP aggregation by small molecules has been considered to be a promising way to eliminate or attenuate hIAPP amyloid associated toxicity.30,31 Consistent with our previous report,17 the present study showed BPA accelerated hIAPP aggregation while seven additional BPs analogues showed different effects on hIAPP amyloid formation (Fig. 8). BPAF, TBBPA and TCBPA showed the highest inhibition abilities, BPB, BPAP exhibited weak effects while BPF and BPS showed no effect on hIAPP aggregation (Fig. 2 and 3). Furthermore, BPAF, BPAP, TBBPA and TCBPA also delayed the transition of hIAPP from unordered structure to β-structures (Fig. 4) and attenuated the membrane disruption effects of hIAPP (Fig. 7).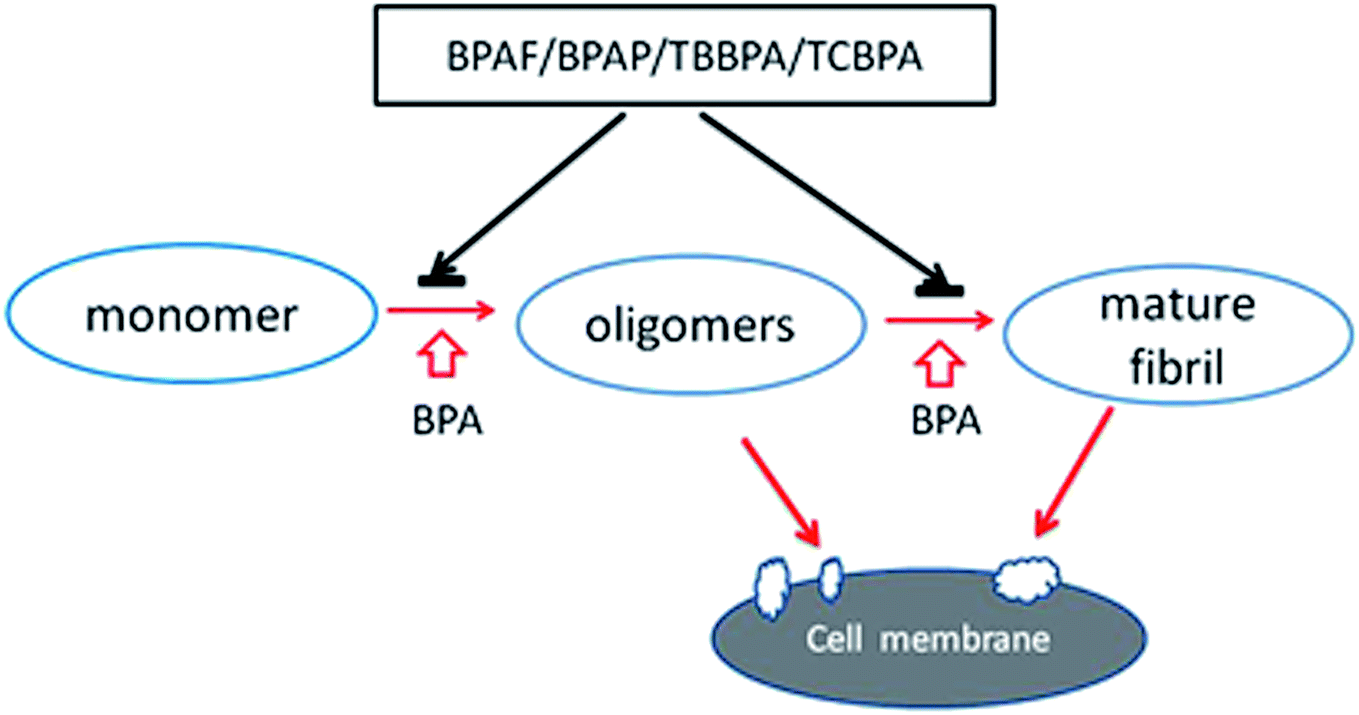 | ||
| Fig. 8 A schematic representation of how BPs analogues affect hIAPP aggregation. BPA promotes the oligomerization and fibrillation of hIAPP, which form pores on membrane which disrupts the membrane. In contrast, BPAF, BPAP, TBBPA and TCBPA inhibit this process and protect the cells. | ||
The strong amyloid inhibitors we identified were BPAF, BPAP, TBBPA and TCBPA, among which, BPAF, TBBPA and TCBPA have halogen substitutions; with the latter two have aromatic halogenation (Fig. 1B). Halogenation has been widely used to provide new properties for organic compounds, such as selective chemical modification of proteins and improved oral absorption/blood–brain barrier permeability of drug candidates.32 Moreover, halogenation of aromatic molecules has been reported to enhance inhibitory capacities of small molecules on Aβ-associated neurotoxicity,32 and single halogen substitutions on the aromatic side-chain of Fmoc-Phe have been shown to affect the subtle balance between steric and electronic effects in π–π interactions and thus enhanced self-assembly.33,34
BPs analogues belong to the polyphenol family, which may competitively interact with aromatic residues in amyloidogenic proteins, prevent the π–π stacking, and block the self-assembly process by inserting between aromatic residues.35,36 The similar chemical structures of BPs analogues but different effects on hIAPP amyloid formation suggest that although the polyaromatic nature may affect amyloid fibril formation by specific aromatic interactions, the atomic substitutions also fine-tune the assembly of hIAPP amyloid.36,37
Endocrine-disrupting compounds (EDCs) have been closely studied recently because they have shown estrogenic effects in fish,24 avian,27 and mammalian cells.25,26 The adverse health effects of exposure to EDCs include decreased sperm count, reduced fertility, and increased incidence of breast, ovarian, and testicular cancers.38,39 BPs analogues studied here have all been listed as EDCs by the United States Environmental Protection Agency (U.S. EPA). The impact of BPs analogues exposure on human health is regarded as an accumulative process due to its widespread penetration in daily life.40 For example, parental exposure to environmentally relevant concentration of BPAF (5 μg L−1) could result in delayed hatching of the offspring of zebrafish.41 TBBPA and TCBPA are reported to show thyroid hormone agonist/antagonist activities42–44 and act as obesogens in zebrafish larvae,45 have greater cytotoxic than BPA in rat hepatocytes.46 Therefore, although our data showed BPAF, BPAP, TBBPA and TCBPA can attenuate the aggregation of hIAPP compare to BPA in vitro, cytotoxicity and haemolysis assays nevertheless suggest their biological safeties awaits further study.
In summary, our data provide the first evidence that four BPs analogues (BPAF, BPAP, TBBPA and TCBPA) can attenuates the aggregation and membrane disruption effect of hIAPP. Our results also suggest that halogenation of polyphenol may increase the inhibitory capacity, with halogenation on the benzene ring more likely show stronger inhibitory capacity than on the side chain. These findings may be helpful for searching or designing novel anti-hIAPP aggregation modulators. Exposure to lipophilic chemicals such as BPA and its analogues has been reported to cause neurological impairments, neurodevelopment disorders and neurodegenerative diseases,47 many of which are associated with amyloidogenic proteins such as amyloid-β and α-synuclein.48 It will also be of great interest to study in the future that how these BPs analogues affect the misfolding of these proteins.
4. Materials and methods
4.1. Materials
hIAPP was purchased from Ketai Biotechnology (Shanghai, China). BPA, BPS, BPAF, BPAP, TBBPA were obtained from Aladdin reagent (Shanghai, China). BPB, BPF, TCBPA were purchased from Tokyo Kasei Kogyo Co., Ltd. (TCI). 2-Oleoyl-1-palmitoyl-sn-glycerol-3-phospho-rac (1-glycerol) sodium salt (POPG), carboxyfluorescein, hexafluoroisopropanol (HFIP) and thioflavin-T (ThT) were purchased from Sigma-Aldrich (St. Louis, USA). All other chemicals were of the highest grade available.4.2. hIAPP aggregation
For amyloid formation, hIAPP was dissolved in 1,1,1,3,3,3-hexafluoroisopropanol (HFIP) and sonicated for 2 min to homogenize the sample as we previously described.49 Then the solution was diluted to desired concentration in 25 mM sodium phosphate buffer (pH 7.4) containing 50 mM NaCl, and a final HFIP concentration of 1%. Freshly prepared BPA, BPB, BPF, BPS, BPAF, BPAP, TBBPA, TCBPA stock solution in DMSO was then immediately added to desired concentrations, thoroughly mixed and ready for further analysis. The preparation process is strictly limited to 5 min.4.3. Thioflavin -T (ThT) fluorescence assay
For aggregation assays, 0.6 μL thioflavin-T (10 mM) was added to 300 μL hIAPP sample (10 μM) in the absence or presence of different compounds. The ThT fluorescence of sample solution in the quartz cuvette was continuously scanned for 2 h quiescently at 25 °C on a Hitachi FL-2700 fluorometer (Hitachi, Tokyo, Japan) to monitor fibril formation. The excitation and emission wavelength were set at 450 nm and 482 nm, respectively. All of the experiments were performed for at least three times.4.4. Transmission electron microscopy (TEM)
TEM was performed as we previously described.49 Briefly, 5 μL hIAPP sample (10 μM) in the absence or presence of different compounds was applied onto a 300-mesh formvar-carbon-coated copper grid (Shanghai, China) followed by staining with 1% fresh prepared uranyl formate. Samples were air-dried and observed under a transmission microscope (Hitachi, Tokyo, Japan) operating at an accelerating voltage of 100 kV.4.5. Circular dichroism (CD)
CD spectra were recorded at 25 °C under a constant flow of N2 by using a JASCO-810 spectropolarimeter (JASCO, Japan). Sample solutions containing 20 μM hIAPP, 25 mM PBS and 50 mM NaCl in the presence or absence of 200 μM of different compounds were incubated in the cuvette at 25 °C for 60 min or 90 min. Data were recorded from 260 to 200 nm with a 1 mm path length, 2 nm bandwidth, 200 nm min−1 scanning speed and 1 s response. The reported spectrum for each sample was the average of at least 3 measurements and the background was subtracted using appropriate controls. The background spectra were obtained using controls containing corresponding concentration of compounds.4.6. Dot blotting
Samples were incubated as we previously described.50 At indicated time points, 4 μL of hIAPP samples (10 μM) in the absence or presence of different compounds were spotted onto nitrocellulose membranes and dried at room temperature. Nitrocellulose membrane was blocked at room temperature for 1 hour with 5% milk and then washed with Tris-buffered saline (TBST, 0.1% Tween 20, 20 mM Tris–HCl, 150 mM NaCl, pH 7.4) for 10 min. Each membrane was then incubated at 4 °C overnight with A11 or OC antibody (1:2500, Millipore, Billerica, USA), which specially recognize oligomer and fibril, respectively. Membranes were further washed in TBST for three times and then incubated with anti-rabbit IgG (1:2000) for 2 h at room temperature. Signal detection was performed using an ECL chemiluminescence substrate kit (Millipore, Billerica, USA).
4.7. Dynamic light scattering (DLS) analysis
DLS was performed by using a zeta pals potential analyzer (Brookhaven Instruments, New York, USA). Solutions containing 10 μM hIAPP, 25 mM PBS and 50 mM NaCl in the presence or absence of 100 μM of different compounds were incubated in the cuvette at 25 °C in the analyzer for 1 h. Compounds alone in 25 mM PBS buffer (containing 50 mM NaCl) were also detected for 24 h. All of the samples were scanned for 3 times (30 s per scan) and the mean particle size and multimodal size distribution were recorded every 5 min.4.8. Dye leakage assays
POPG was dissolved in a mixture of chloroform/methanol (1:1) at a concentration of 10 mg mL−1, and then the solvent was removed under a stream of N2 and freeze drying overnight. The dried POPG film was mixed with 25 mM PBS (50 mM NaCl, pH 7.4) containing 40 mM carboxyfluorescein to prepare multilamellar vesicles. PD-10 desalting column (Sangon, Shanghai, China) was used to remove non-encapsulated carboxyfluorescein. 1% (v/v) POPG vesicles encapsulating carboxyfluorescein in 25 mM PBS (50 mM NaCl, pH 7.4) were used for fluorescence measurements with excitation and emission wavelength set at 493 nm and 518 nm, respectively.51 Prepared hIAPP samples were added to POPG vesicles at a final concentration of 1 μM immediately before measurement. POPG vesicles alone were tested as baseline and the signals of POPG vesicles treated with 0.2% (v/v) Triton-X 100 (complete membrane leakage) were measured as positive control. All of the experiments were performed at least three times.
4.9. MTT cell toxicity assay
MTT-based cell toxicity assays were performed as we previously described.52 BxPC3 cells, a human pancreatic tumor cell line, were cultured in DMEM high glucose medium containing 10% FBS, 1% penicillin-streptomycin solution and 1% sodium pyruvate. Cells were plated in 96-well plates at a density of 5 × 103 cells per well and cultured for 24 h. The medium was then replaced with fresh medium containing hIAPP (1 μM) and varied amounts of BPA analogues for 24 h further incubation (containing 1.1% DMSO). Cells treated with BPA analogues were used as the controls. For MTT assay, cells were co-incubated with 10 μL MTT (5 mg mL−1) per well for 3 h. 100 μL formazan buffer was then added to each well and the absorbance was measured at 570 nm.4.10. Haemolysis assay
Haemolysis assay was performed as we previously described.53 Fresh blood was centrifuged at 1000g for 10 min, and erythrocytes were separated from plasma and washed three times with isotonic phosphate buffered saline (pH 7.4). For haemolytic assay, cell suspensions (1% haematocrit) were incubated at 37 °C for 12 h in the presence or absence of BPA analogues of hIAPP under pH 7.4 with a concentration of 10 μM. Absorbance of the supernatant was determined at 540 nm. The haemolytic rate was calculated in relation to the haemolysis of erythrocytes in 10 mM phosphate buffer, which was taken as 100%.4.11. Aggregator Advisor prediction
Aggregator Advisor (http://advisor.bkslab.org/) is used as described21 to predict molecules that aggregate or may aggregate under biochemical assay conditions, which is based on the chemical similarity to known aggregators, and physical properties.4.12. Statistical analysis
All results were expressed as the mean ± SD. The Kruskal–Wallis test and the Mann–Whitney test were used to assess statistical significance. Difference was considered statistically significant at P < 0.05.Author contributions
K. H. conceived the ideal and designed the experiments; L. H., M. L., Y. Z., X. Y., L. M., H. G. performed study; L. H, M. L., K. H. wrote the paper.Conflict of interest
The authors declare no conflict of interest regarding this work.Abbreviations
| hIAPP | Human islet amyloid polypeptide |
| T2DM | Type 2 diabetes mellitus |
| BPs | Bisphenols |
| BPA | Bisphenol A |
| BPB | Bisphenol B |
| BPF | Bisphenol F |
| BPS | Bisphenol S |
| BPAF | Bisphenol AF |
| BPAP | Bisphenol AP |
| TBBPA | Tetrabromobisphenol A |
| TCBPA | Tetrachlorobisphenol A |
| Aβ | β-Amyloid |
| AD | Alzheimer's disease |
| POPG | 2-Oleoyl-1-palmitoyl-sn-glycerol-3-phospho-rac (1-glycerol) sodium salt |
| HFIP | Hexafluoroisopropanol |
| ThT | Thioflavin-T |
| TEM | Transmission electron microscopy |
| DLS | Dynamic light scattering |
| CD | Circular dichroism |
| TBST | Tris-buffered saline |
Acknowledgements
This work was supported by the Natural Science Foundation of China (No. 81172971, 81222043 and 31471208), the Municipal Key Technology Program of Wuhan (Wuhan Bureau of Science & Technology, No. 201260523174) and the Natural Science Foundation of Hubei Province (2014CF021). BxPC3 cell line is a kind gift of Dr Ming Xiang (HUST).References
- C. L. Masters, G. Simms, N. A. Weinman, G. Multhaup, B. L. McDonald and K. Beyreuther, Proc. Natl. Acad. Sci. U. S. A., 1985, 82, 4245–4249 CrossRef CAS.
- G. G. Glenner and C. W. Wong, Biochem. Biophys. Res. Commun., 1984, 120, 885–890 CrossRef CAS PubMed.
- M. Baba, S. Nakajo, P. H. Tu, T. Tomita, K. Nakaya, V. M. Y. Lee, J. Q. Trojanowski and T. Iwatsubo, Am. J. Pathol., 1998, 152, 879–884 CAS.
- P. Westermark and L. Grimelius, Acta Pathol. Microbiol. Scand., Sect. A, 1973, 81, 291–300 CAS.
- A. Clark, G. J. Cooper, C. E. Lewis, J. F. Morris, A. C. Willis, K. B. Reid and R. C. Turner, Lancet, 1987, 2, 231–234 CrossRef CAS.
- R. L. Hull, G. T. Westermark, P. Westermark and S. E. Kahn, J. Clin. Endocrinol. Metab., 2004, 89, 3629–3643 CrossRef CAS PubMed.
- M. Stefani and C. M. Dobson, J. Mol. Med., 2003, 81, 678–699 CrossRef CAS PubMed.
- L. A. Scrocchi, Y. Chen, S. Waschuk, F. Wang, S. Cheung, A. A. Darabie, J. McLaurin and P. E. Fraser, J. Mol. Biol., 2002, 318, 697–706 CrossRef CAS PubMed.
- J. Jayamani, G. Shanmugam and E. R. A. Singam, RSC Adv., 2014, 4, 62326–62336 RSC.
- T. Geens, L. Goeyens and A. Covaci, Int. J. Hyg. Environ. Health, 2011, 214, 339–347 CrossRef CAS PubMed.
- S. De Coster and N. van Larebeke, J. Environ. Public Health, 2012, 2012, 713696 Search PubMed.
- C. Y. Liao, F. Liu and K. Kannan, Environ. Sci. Technol., 2012, 46, 6515–6522 CrossRef CAS PubMed.
- L. Grumetto, D. Montesano, S. Seccia, S. Albrizio and F. Barbato, J. Agric. Food Chem., 2008, 56, 10633–10637 CrossRef CAS PubMed.
- C. A. de Wit, D. Herzke and K. Vorkamp, Sci. Total Environ., 2010, 408, 2885–2918 CrossRef CAS PubMed.
- H. Gong, X. Zhang, B. Cheng, Y. Sun, C. Li, T. Li, L. Zheng and K. Huang, PLoS One, 2013, 8, e54198 CAS.
- H. LeVine 3rd, Protein Sci., 1993, 2, 404–410 CrossRef CAS PubMed.
- L. Jiao, X. Zhang, L. Huang, H. Gong, B. Cheng, Y. Sun, Y. Li, Q. Liu, L. Zheng and K. Huang, Food Chem. Toxicol., 2013, 56, 398–405 CrossRef CAS PubMed.
- C. E. Higham, E. T. A. S. Jaikaran, P. E. Fraser, M. Gross and A. Clark, FEBS Lett., 2000, 470, 55–60 CrossRef CAS PubMed.
- A. R. A. Ladiwala, J. C. Lin, S. S. Bale, A. M. Marcelino-Cruz, M. Bhattacharya, J. S. Dordick and P. M. Tessier, J. Biol. Chem., 2010, 285, 24228–24237 CrossRef CAS PubMed.
- R. Kayed, E. Head, F. Sarsoza, T. Saing, C. W. Cotman, M. Necula, L. Margol, J. Wu, L. Breydo, J. L. Thompson, S. Rasool, T. Gurlo, P. Butler and C. G. Glabe, Mol. Neurodegener., 2007, 2, 18 CrossRef PubMed.
- J. J. Irwin, D. Duan, H. Torosyan, A. K. Doak, K. T. Ziebart, T. Sterling, G. Tumanian and B. K. Shoichet, J. Med. Chem., 2015, 58, 7076–7087 CrossRef CAS PubMed.
- N. B. Last, E. Rhoades and A. D. Miranker, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 9460–9465 CrossRef CAS PubMed.
- A. A. Meratan, A. Ghasemi and M. Nemat-Gorgani, J. Mol. Biol., 2011, 409, 826–838 CrossRef CAS PubMed.
- B. P. Baldigo, S. D. George, P. J. Phillips, J. D. C. Hemming, N. D. Denslow and K. J. Kroll, Environ. Toxicol. Chem., 2015, 34, 2803–2815 CrossRef CAS PubMed.
- J. W. Finger and R. M. Gogal, Arch. Environ. Contam. Toxicol., 2013, 65, 704–714 CrossRef CAS PubMed.
- Y. Li, C. J. Luh, K. A. Burns, Y. Arao, Z. L. Jiang, C. T. Teng, R. R. Tice and K. S. Korach, Environ. Health Perspect., 2013, 121, 459–466 Search PubMed.
- M. A. Ottinger, E. Lavoie, N. Thompson, A. Barton, K. Whitehouse, M. Barton, M. Abdelnabi, M. Quinn Jr, G. Panzica and C. Viglietti-Panzica, Brain Res. Rev., 2008, 57, 376–385 CrossRef CAS PubMed.
- R. L. Hull, G. T. Westermark, P. Westermark and S. E. Kahn, J. Clin. Endocrinol. Metab., 2004, 89, 3629–3643 CrossRef CAS PubMed.
- S. Costes, R. Langen, T. Gurlo, A. V. Matveyenko and P. C. Butler, Diabetes, 2013, 62, 327–335 CrossRef CAS PubMed.
- L. Q. Huang, X. R. Liu, B. A. Cheng and K. Huang, Arch. Biochem. Biophys., 2015, 568, 46–55 CrossRef CAS PubMed.
- B. Cheng, H. Gong, H. W. Xiao, R. B. Petersen, L. Zheng and K. Huang, Biochim. Biophys. Acta, Gen. Subj., 2013, 1830, 4860–4871 CrossRef CAS PubMed.
- M. Z. Hernandes, S. M. Cavalcanti, D. R. Moreira, W. F. de Azevedo Junior and A. C. Leite, Curr. Drug Targets, 2010, 11, 303–314 CrossRef CAS PubMed.
- D. M. Ryan, S. B. Anderson and B. L. Nilsson, Soft Matter, 2010, 6, 3220–3231 RSC.
- H. E. Wong, J. A. Irwin and I. Kwon, PLoS One, 2013, 8, e57288 CAS.
- E. Ahmad, A. Ahmad, S. Singh, M. Arshad, A. H. Khan and R. H. Khan, Biochimie, 2011, 93, 793–805 CrossRef CAS PubMed.
- Y. Porat, A. Abramowitz and E. Gazit, Chem. Biol. Drug Des., 2006, 67, 27–37 CAS.
- M. Levy, Y. Porat, E. Bacharach, D. E. Shalev and E. Gazit, Biochemistry, 2008, 47, 5896–5904 CrossRef CAS PubMed.
- F. S. vom Saal, S. C. Nagel, P. Palanza, M. Boechler, S. Parmigiani and W. V. Welshons, Toxicol. Lett., 1995, 77, 343–350 CrossRef CAS PubMed.
- Y. Takai, O. Tsutsumi, Y. Ikezuki, H. Hiroi, Y. Osuga, M. Momoeda, T. Yano and Y. Taketani, Biochem. Biophys. Res. Commun., 2000, 270, 918–921 CrossRef CAS PubMed.
- J. S. Lakind and D. Q. Naiman, J. Exposure Sci. Environ. Epidemiol., 2008, 18, 608–615 CrossRef CAS PubMed.
- J. C. Shi, Z. H. Jiao, S. Zheng, M. Li, J. Zhang, Y. X. Feng, J. Yin and B. Shao, Chemosphere, 2015, 128, 252–257 CrossRef CAS PubMed.
- M. Terasaki, K. Kosaka, S. Kunikane, M. Makino and F. Shiraishi, Chemosphere, 2011, 84, 1527–1530 CrossRef CAS PubMed.
- H. Sun, O. X. Shen, X. R. Wang, L. Zhou, S. Q. Zhen and X. D. Chen, Toxicol. in Vitro, 2009, 23, 950–954 CrossRef CAS PubMed.
- S. Kitamura, T. Kato, M. Iida, N. Jinno, T. Suzuki, S. Ohta, N. Fujimoto, H. Hanada, K. Kashiwagi and A. Kashiwagi, Life Sci., 2005, 76, 1589–1601 CrossRef CAS PubMed.
- A. Riu, C. W. McCollum, C. L. Pinto, M. Grimaldi, A. Hillenweck, E. Perdu, D. Zalko, L. Bernard, V. Laudet, P. Balaguer, M. Bondesson and J. A. Gustafsson, Toxicol. Sci., 2014, 139, 48–58 CrossRef CAS PubMed.
- Y. Nakagawa, T. Suzuki, H. Ishii and A. Ogata, Xenobiotica, 2007, 37, 693–708 CrossRef CAS PubMed.
- H. I. Zeliger, Interdiscip. Toxicol., 2013, 6, 103–110 Search PubMed.
- E. Zerovnik, Eur. J. Biochem., 2002, 269, 3362–3371 CrossRef CAS PubMed.
- X. Zhang, B. Cheng, H. Gong, C. Li, H. Chen, L. Zheng and K. Huang, FEBS Lett., 2011, 585, 71–77 CrossRef CAS PubMed.
- H. Gong, Z. He, A. Peng, X. Zhang, B. Cheng, Y. Sun, L. Zheng and K. Huang, Sci. Rep., 2014, 4, 5648 CAS.
- J. R. Brender, E. L. Lee, M. A. Cavitt, A. Gafni, D. G. Steel and A. Ramamoorthy, J. Am. Chem. Soc., 2008, 130, 6424–6429 CrossRef CAS PubMed.
- B. Cheng, H. Gong, X. Li, Y. Sun, X. Zhang, H. Chen, X. Liu, L. Zheng and K. Huang, Biochem. Biophys. Res. Commun., 2012, 419, 495–499 CrossRef CAS PubMed.
- Y. Li, L. Huang, X. Yang, C. Wang, Y. Sun, H. Gong, Y. Liu, L. Zheng and K. Huang, Biochem. Biophys. Res. Commun., 2013, 440, 56–61 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra21792j |
| This journal is © The Royal Society of Chemistry 2016 |