
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Chloride anion transporters inhibit growth of methicillin-resistant Staphylococcus aureus (MRSA) in vitro†
Andrew I.
Share
a,
Khushali
Patel
a,
Cristina
Nativi
b,
Eun J.
Cho
c,
Oscar
Francesconi
*b,
Nathalie
Busschaert‡
d,
Philip A.
Gale
 *d,
Stefano
Roelens
*b and
Jonathan L.
Sessler
*a
*d,
Stefano
Roelens
*b and
Jonathan L.
Sessler
*a
aDepartment of Chemistry, The University of Texas, Austin, TX 78712-1224, USA. E-mail: sessler@cm.utexas.edu
bDepartment of Chemistry and INSTM, University of Florence, Polo Scientifco e Tecnologico, 50019 Sesto Fiorentino, Firenze, Italy. E-mail: oscar.francesconi@unifi.it; stefano.roelens@unifi.it
cTargeted Therapeutic Drug Discovery and Development (TTDDDP), College of Pharmacy, University of Texas-Austin, Austin, TX 78723, USA
dChemistry, University of Southampton, Southampton, SO17 1BJ, UK. E-mail: philip.gale@soton.ac.uk
First published on 18th May 2016
Abstract
A series of aminopyrrolic receptors were tested as anion transporters using POPC liposome model membranes. Many were found to be effective Cl− transporters and to inhibit clinical strains of Staphylococcus aureus growth in vitro. The best transporters proved effective against the methicillin-resistant Staphylococcus aureus (MRSA) strains, Mu50 and HP1173. Tris-thiourea tren-based chloride transporters were also shown to inhibit the growth of S. aureus in vitro.
There is a tremendous interest in novel antibiotics that can combat highly resistant bacterial strains such as MRSA.1 Recent studies of compounds that can mediate the transport of anions demonstrated significant biological activity. Several have shown antiproliferative activity in cancer cell lines.2,3 For instance, the strapped calixpyrrole (1) displays modest NaCl transport in liposome models and is able to inhibit growth of cancer cell lines in vitro.4 This effect was ascribed to a combination of receptor mediated chloride transport and sodium transport involving endogenous ion channels. This and other recent developments in the anion transport field5,6 have lead us to consider that other biologically active species may be mediating their effect in whole or in part by an ability to transport chloride anions7 into cells.§¶
As a first test of this hypothesis, two known antibiotic agents, thiocarlide and trichlorocarbanalide were tested for their ability to transport chloride anions through a standard 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomal model membrane. They were found to be modest NaCl (or HCl||) co-transporters and effective Cl−/NO3− antiporters (Fig. 1). This finding, as well as the ability of several cation ionophores to act as antibiotics,8 has led us to consider that anion transporters might show antibiotic activity.

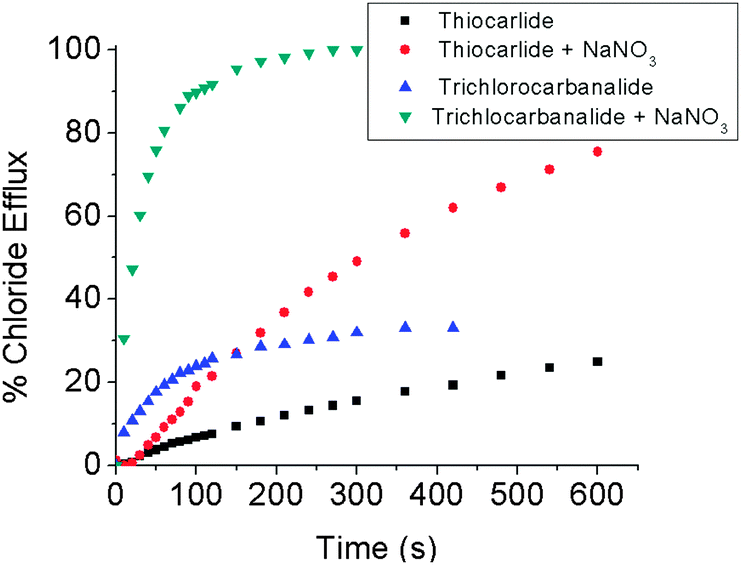 | ||
| Fig. 1 Chloride efflux caused by the addition of the antibiotics thiocarlide or trichlorocarbanalide (0.675 mol%), with and without NaNO3 added to the exterior of POPC vesicles loaded with 0.5 M NaCl, 0.1 M Na2SO4, and 20 mM phosphate buffer, pH = 7.2. 100% chloride efflux was determined by lysing the liposomes with Triton X. | ||
With this view in mind, we selected a family of aminopyrrolic compounds, namely 2–15 (Scheme 1), and investigated their chloride anion transport properties. The antibiotic activity of these compounds, as well as the known Cl−/HCO3− anion antiport agents 16–18 (Scheme 2), were also tested. The present study was prompted by previous work, showing that several aminopyrrolic compounds were biologically active vs. pathogenic yeasts,10 exhibited antiviral activity,11 induced apoptosis in HeLa and PLC/PRF/5 cells,12 and were found to recognize the chloride anion well in polar organic solvents.13,14 Here, we report the finding that for the aminopyrrolic compounds 2–15 chloride anion transport in liposomal model correlates well with in vitro antibiotic activity against the S. aureus strains UAMS1, HP1173, and Mu50.
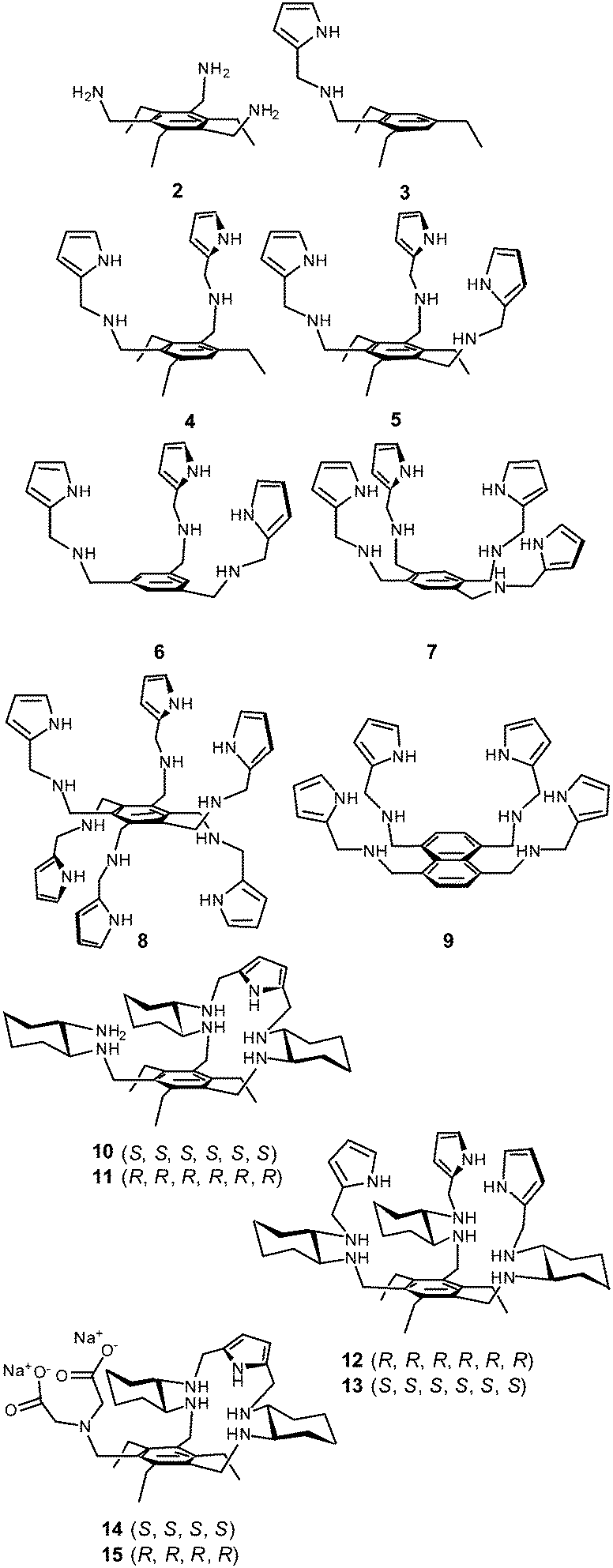 | ||
| Scheme 1 Structures of compounds 2–15. | ||
 | ||
| Scheme 2 Structures of compounds 16–18. | ||
Compounds 2–15 were examined for their ability to transport chloride anions across a phospholipid bilayer. In a typical experiment, 160 nm POPC vesicles that contained NaCl (500 mM), Na2SO4 (100 mM), and HEPES buffer (5 mM) were suspended in a solution of Na2SO4 (100 mM) and HEPES buffer (5 mM). The compound subject to study (0.675 mol%, compared to POPC concentration) was added to the solution and the rate of chloride efflux from the vesicles was monitored using a Cl− ion selective electrode (ISE). After 9 minutes, the liposomes were lysed with polyethylene glycol tert-octylphenyl ether (Triton X-114), and a final reading from the ISE corresponding to 100% chloride release was taken. High levels of transport activity were seen in descending order for compounds 12, 13, 10, 8, and 11 (Fig. 2). In these studies the solutions external to the liposomes contained only SO42− and HEPES. These are hydrophilic species15 that are typically not transported through POPC membranes.16 Therefore, we infer that under these experimental conditions, anion antiport should not contribute to the efflux of chloride from the liposome. Studies involving compound 5 were also carried out when the external sulphate anion was replaced by either NO3− or HCO3− (as the sodium salts). The nitrate and bicarbonate anions are considerably more lipophilic than the sulphate anion and can facilitate the transport of chloride through the liposomal membrane by promoting Cl−/NO3− or Cl−/HCO3− antiport processes. In these experiments a higher concentration of NaCl (1 M) inside the liposome was used to give a larger response with the ISE. The rate of chloride efflux is slightly raised on the addition of the more lipophilic anions nitrate and bicarbonate to the external solution (Fig. 3). Nevertheless, this difference only accounts for a small percentage of the total efflux of chloride from the liposomes. We thus conclude that anion antiport only plays a minor role in chloride efflux and that the receptors of the present study are functioning primarily as cation–anion cotransporters.||
 | ||
| Fig. 2 Chloride efflux was initiated by the addition of 2–15 (0.675 mol%) to a solution of POPC vesicles (1 mM POPC, internal solution: 0.5 M NaCl, 0.1 M Na2SO4, 5 mM HEPES, pH = 7.2, external solution: 0.1 M Na2SO4, 5 mM HEPES pH = 7.2). | ||
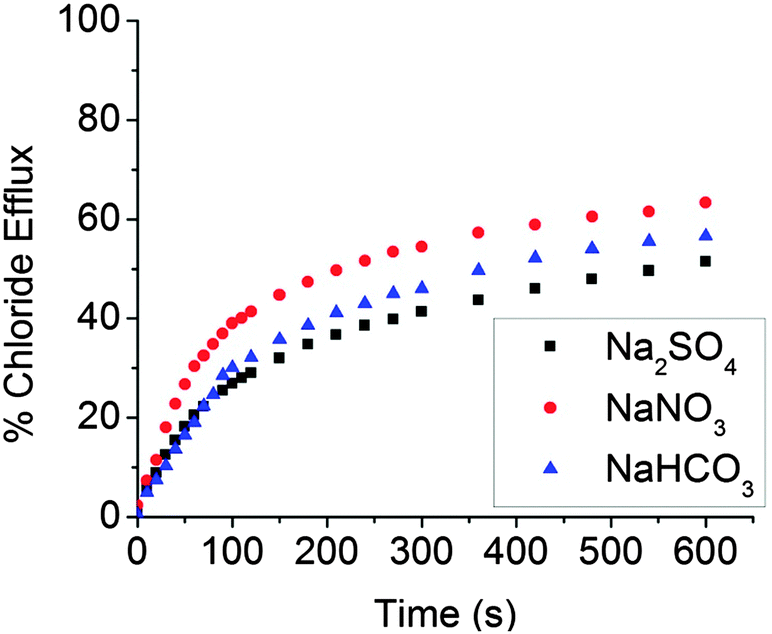 | ||
| Fig. 3 Chloride efflux promoted by the addition of 5 (0.675 mol%) and either NaNO3 or NaHCO3 to the liposome solution (1 mM POPC, internal solution: 1 M NaCl, 0.1 M Na2SO4 5 mM HEPES, pH = 7.2, external solution: 0.1 M Na2SO4, 5 mM HEPES pH = 7.2). | ||
Studies of the antibiotic activity of compounds 2–18 were carried out on 96 well plates using the broth dilution method (cf.Fig. 4). Antibiotic activity was tested against both Gram positive (S. aureus) and Gram negative (Pseudomonus aeruginosa) bacteria. The bacterial laden plates were incubated for 16 hours at 37 °C. The plates were then examined visually. The wells that remained clear were deemed to reflect bacterial growth inhibition. Wells that were cloudy were considered to contain active colonies of bacteria. The MIC was taken as the concentration present in the last well that remained clear as judged in this manner. Experiments were repeated 3 times and gave concordant results within the range of a single serial dilution (Tables S1–S5, ESI†). Good activity against S. aureus was seen for many compounds. However, except for compounds 5 and 8, no activity was seen against Gram negative bacteria.** The most active aminopyrrole compounds proved to be in descending order 5, 8, 10, 11, 12, 13. The lowest activity was found for 2, 14, and 15 (Table 1). A plot of MIC vs. chloride anion transport revealed a general correlation with bacterial inhibition in S. aureus (Fig. 5). This trend was found to hold best for the compounds with intermediate activity.
 | ||
| Fig. 4 A 96 well plate bacterial assay of compounds 18 (top row) and 17 (middle row) against S. aureus grown in 99 μL brain–heart infusion media and DMSO (bottom row) after 16 hours of bacterial growth. Each successive well (from left to right) represents a two-fold dilution of compound. Clear wells indicate inhibition of bacterial growth, while cloudy wells signify unhindered bacterial growth. | ||
| Compound | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC (μg mL−1) | >100 | 50 | 66.67 | 6.25 | 25 | 50 | 4.69 | 25 | 12.5 | 12.5 | 4.69 | 3.13 | >100 | >100 |
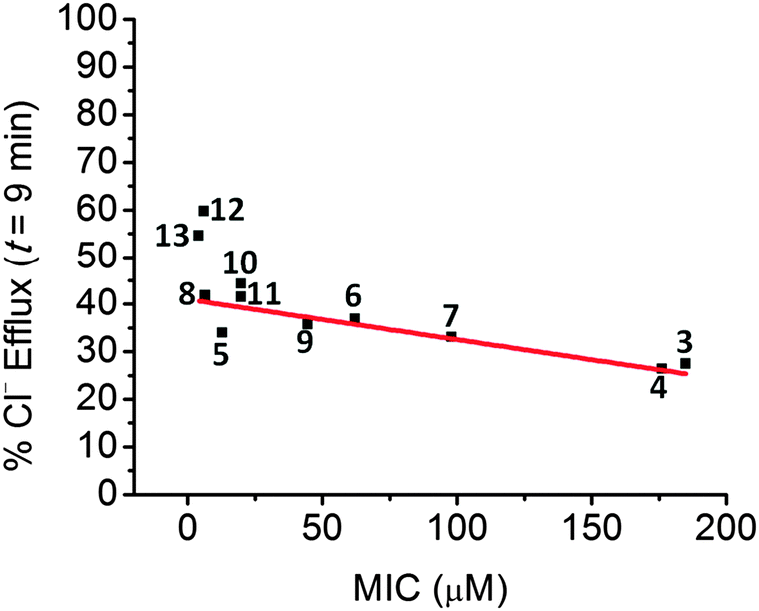 | ||
| Fig. 5 Plot of chloride efflux promoted by 2–13 in liposomal transport models as detailed in Fig. 2 against their effectiveness in reducing the growth of the Mu50 (resistant) strain of S. aureus. | ||
While the ion transport rates correlate with the antibiotic activity of the compounds, the deviations of the most active compounds may reflect the fact that ion transport ability is only part of the story underlying the observed antibiotic activity. In particular, the best chloride anion transporters, 12 and 13 have significantly higher transport rates than compounds 10 and 11, but only modestly better antibacterial activity. Previous studies of these compounds revealed that compounds 12 and 13 can bind to mannosides on the cell glycocalix,10 and that this may underlie their observed in vitro anticancer activity, as opposed to anion transport per se. Such findings provide support for the notion that the antibiotic activity of 12 and 13 may be due to the compounds acting by multiple mechanisms. Additionally, compounds 5 and 8 were the only compounds to be active against Gram negative bacteria. This leads to the conclusion that, at least in the case of compounds 5, 8, 12, and 13, ion transport is not the only factor responsible for antibiotic activity.
Nevertheless, we believe that an ability to promote the through-membrane transport of the chloride anion may be a useful harbinger of antibiotic activity. The fact that chloride anion exchange, rather than cation–anion cotransport, was seen for the known antibiotics, thiocarlide and trichlorocarbanalide (cf.Fig. 1), leads us to suggest that chloride anion transport can inhibit bacterial growth in vitro. Further evidence for this suggestion came from the finding that the synthetic thioureas 16–18 proved quite active in vitro (MIC = 1.23, 0.93, and 1.78 μg mL−1 for the Mu50 S. aureus, respectively). These species are known to act primarily as anion exchangers (e.g., Cl− for HCO3− and Cl− for SO42−), rather than as ion pair cotransporters,2,16 although this class of transporter has been shown to be capable of dissipating pH gradients across lipid bilayers.2,17
A notable feature of the present compounds is that they are active against HP1173 and Mu50 methicillin resistant strains of S. aureus, as well as the non-methicillin resistant UAMS1 strain. This leads us to suggest that receptors that can act as highly effective through-membrane anion carriers (both exchangers and ion pair cotransporters) may have a role to play as rationally designed antibiotic agents. Nevertheless, it is important to appreciate that the correlation between chloride anion transport and observed antibiotic activity found for 2–15 in the case of S. aureus is not perfect. While a general trend holds, deviations are seen in the case of the most active compounds. This may indicate that there are other mechanisms aside from ion transport that are responsible for the antibacterial activity in S. aureus or that there are differences between the properties of the model membranes used in this study and the bacterial membrane.18 Further study of these and other anion transporters are on-going in an effort to elucidate more fully the underlying mechanisms of action and to develop more active receptor-based antibiotic agents.
The work in Austin was supported by the National Institutes of Health (grant no. GM 058907 and GM 103790) and the Robert A. Welch Foundation (grant F-1018 to J. L. S.). Financial support from Ente Cassa di Risparmio di Firenze (ECR) and Cassa di Risparmio Firenze (CR Firenze) for the work in Florence is gratefully acknowledged. N. B. thanks the University of Southampton and the A*STAR ARAP programme for a PhD studentship. P. A. G. thanks the Royal Society and the Wolfson Foundation for a Research Merit Award and the EPSRC (EP/K039466/1) (Core Capability for Chemistry Research in Southampton). We thank Dr Aimee Wessler and Prof. Marvin Whiteley for help with the antibacterial cell studies.
Notes and references
- M. E. Stryjewski and G. R. Corey, Clin. Infect. Dis., 2014, 58, S10–S19 CrossRef CAS PubMed.
- N. Busschaert, M. Wenzel, M. E. Light, P. Iglesias-Hernández, R. Pérez-Tomás and P. A. Gale, J. Am. Chem. Soc., 2011, 133, 14136–14148 CrossRef CAS PubMed.
- (a) S. J. Moore, M. Wenzel, M. E. Light, R. Morley, S. J. Bradberry, P. Gómez-Iglesias, V. Soto-Cerrato, R. Pérez-Tomás and P. A. Gale, Chem. Sci., 2012, 3, 2501–2509 RSC; (b) S. J. Moore, C. J. E. Haynes, J. González, J. L. Sutton, S. J. Brooks, M. E. Light, J. Herniman, G. J. Langley, V. S. Cerrato, R. Pérez-Tomás, I. Marques, P. J. Costa, V. Félix and P. A. Gale, Chem. Sci., 2013, 4, 103–117 RSC.
- S.-K. Ko, S. K. Kim, A. Share, V. M. Lynch, J. Park, W. Namkung, W. V. Rossom, N. Busschaert, P. A. Gale, J. L. Sessler and I. Shin, Nat. Chem., 2014, 6, 885–892 CrossRef CAS PubMed.
- N. Busschaert and P. A. Gale, Angew. Chem., Int. Ed., 2013, 52, 1374–1382 CrossRef CAS PubMed.
- A. V. Jentzsch, A. Henning, J. Mareda and S. Matile, Acc. Chem. Res., 2013, 46, 2791–2800 CrossRef PubMed.
- (a) H. Li, H. Valkenier, L. W. Judd, P. R. Brotherhood, S. Hussain, J. A. Cooper, O. Jurček, H. A. Sparkes, D. N. Sheppard and A. P. Davis, Nat. Chem., 2014, 8, 24–32 CrossRef PubMed; (b) J. T. Davis, O. Okunola and R. Quesada, Chem. Soc. Rev., 2010, 39, 3843–3862 RSC; (c) G. W. Gokel and S. Negin, Acc. Chem. Res., 2013, 46, 2824–2833 CrossRef CAS PubMed.
- D. A. Kevin II, D. A. F. Meujo and M. T. Hamann, Expert Opin. Drug Discovery, 2009, 4, 109–146 CrossRef PubMed.
- C. R. Elie, G. David and A. R. Schmitzer, J. Med. Chem., 2015, 58, 2358–2366 CrossRef CAS PubMed.
- C. Nativi, O. Francesconi, G. Gabrielli, I. De Simone, B. Turchetti, T. Mello, L. D. C. Mannelli, C. Ghelardini, P. Buzzini and S. Roelens, Chem. – Eur. J., 2012, 18, 5064–5072 CrossRef CAS PubMed.
- O. Francesconi, C. Nativi, G. Gabrielli, I. De Simone, S. Noppen, J. Balzarini, S. Liekens and S. Roelens, Chem. – Eur. J., 2015, 21, 10089–10093 CrossRef CAS PubMed.
- S.-H. Park, Y. P. Choi, J. Park, A. Share, O. Francesconi, C. Nativi, W. Namkung, J. L. Sessler, S. Roelens and I. Shin, Chem. Sci., 2015, 6, 7284–7292 RSC.
- S. Roelens, A. Vacca and C. Venturi, Chem. – Eur. J., 2009, 15, 2635–2644 CrossRef CAS PubMed.
- S. Roelens, A. Vacca, O. Francesconi and C. Venturi, Chem. – Eur. J., 2009, 15, 8296–8302 CrossRef CAS PubMed.
- Y. Marcus, J. Chem. Soc., Faraday Trans., 1991, 87, 2995–2999 RSC.
- N. Busschaert, L. E. Karagiannidis, M. Wenzel, C. J. E. Haynes, N. J. Wells, P. G. Young, D. Makuc, J. Plavec, K. A. Jolliffe and P. A. Gale, Chem. Sci., 2014, 5, 1118–1127 RSC.
- X. Wu, L. W. Judd, E. N. W. Howe, A. M. Withecombe, V. Soto-Cerrato, H. Li, N. Busschaert, H. Valkenier, R. Pérez-Tomás, D. N. Sheppard, Y. Jiang, A. P. Davis and P. A. Gale, Chem, 2016 DOI:10.1016/j.chempr.2016.04.002.
- M. J. Spooner and P. A. Gale, Chem. Commun., 2015, 51, 4883–4886 RSC.
Footnotes |
| † Electronic supplementary information (ESI) available: Details of transport and antibiotic studies. See DOI: 10.1039/c6cc03645g |
| ‡ Present address: Silver Center for Arts and Science, New York University, Department of Chemistry, 100 Washington Square E, New York, NY 10003, USA. |
| § Simple cation transporters often function as antibiotics.8 |
| ¶ During the course of this work, Schmitzer and coworkers reported that certain imidazolium-based anion transporters likewise displayed strong antibiotic effects.9 |
| || The counter cation accompanying chloride anion egress was not monitored directly. The observation of a cation dependence on the rate when the inner solution was changed from NaCl to KCl so as to provide a less hydrated cation14 is consistent with a Na+/Cl− cotransport mechanism (see Supporting Information). However, the presence of readily protonated amino functionality in receptors 2–15 makes it difficult to discount an alternative mechanism involving H+/Cl− cotransport. Through-membrane proton transport is expected to be most important at early times before a proton gradient is built up across the liposomal membrane. An effort to distinguish between limiting NaCl vs. HCl cotransport mechanisms has not been made in the case of thiocarlide and trichlorocarbanalide. |
| ** Although further study is needed, it is likely that artificial receptors such as those of the present study will prove more effective as both ion carriers and antibiotics in the case of Gram positive bacteria than Gram negative bacteria since Gram positive bacteria have a single cellular membrane, whereas Gram negative bacteria have 2 cell membranes. |
| This journal is © The Royal Society of Chemistry 2016 |