Perfluoroalkyl compounds (PFCs) in wildlife from an urban estuary†
Margaret D.
Sedlak
*a and
Denise J.
Greig
b
aSan Francisco Estuary Institute, 4911 Central Avenue, Richmond, CA 94804. E-mail: meg@sfei.org
bThe Marine Mammal Center, 2000 Bunker Road, Fort Cronkhite, Sausalito, CA 94965
First published on 2nd December 2011
Abstract
Previous research has documented the bioaccumulation of perfluoroalkyl compounds (PFCs) in apex predators in remote locations but few studies have evaluated urban estuaries. To assess the importance of PFCs in San Francisco Bay, two apex predators in the San Francisco Bay, double-crested cormorants (Phalacrocorax auritus) and Pacific harbor seals (Phoca vitulina richardii), were sampled. Prey fish (Atherinops affinis and Menidia audens) were also evaluated to better understand potential sources of PFCs to the foodweb. Perfluorooctane sulfonate (PFOS) was the primary PFC detected in cormorant eggs, small fish and harbor seal serum. PFOS detected in San Francisco Bay seal serum was typically an order of magnitude higher than those at the reference site. PFOS concentrations were highest in seals and cormorant eggs from the highly urbanized southern portion of the Bay. PFOS in eggs from the southern part of the Bay remained relatively constant between 2006 and 2009 despite the phase-out of perfluorosulfonyl-based compounds nationally. In addition, these levels exceed the avian predicted no effects concentration of 1.0 μg mL−1. Concentrations of the remaining PFCs measured were substantially lower than those of PFOS.
Environmental impactThis project evaluated concentrations of perfluoroalkyl compounds (PFCs) in apex predators residing in San Francisco Bay (i.e., Double-crested cormorants (Phalacrocorax auritus) and Pacific harbor seals (Phoca vitulina richardii)). Prey fish (Atherinops affinis and Menidia audens) were also monitored to evaluate food web dynamics. Perfluorooctane sulfonate (PFOS) was detected in all tissues; in some instances, at quite high concentrations. In seal serum, concentrations in San Francisco Bay were typically an order of magnitude higher than those measured at the reference site. In general, higher concentrations were observed in animals residing the southern portion of the estuary. PFOS concentrations in eggs from this area exceeded the avian predicted no effects concentration of 1.0 μg mL−1. |
Introduction
Since their initial production in the late 1940s, perfluoroalkyl compounds (PFCs) have been used extensively in a number of industrial, commercial, and consumer applications such as stain and water repellants for carpets, textiles, and paper products, insecticides, aqueous film-forming foams, surfactants, fluoropolymer production, metal-plating and cleaning, and lubricants.1 PFCs are widely used because they are chemically and thermally stable, have excellent surface tension lowering properties, and are both hydrophobic and oleophobic.Perfluorooctane sulfonate (PFOS) and to a lesser extent, eight to 12 carbon chain perfluorocarboxylates (C8–C12 PFCAs) have been widely detected in biota around the world including marine mammals, birds, fish, reptiles, and benthic organisms.2,3 The sources of these compounds are likely from industrial, commercial and consumer use of the PFC-containing products and from the abiotic and biotic transformation of PFC precursors. PFOS and PFCAs are the terminal transformation products of a number of precursors such as perfluorooctansulfonamides,4 and perfluoroalkyl sulfonamide alcohols5 and, for PFCAs, fluorotelomer alcohols.6,7 The long range transport of these volatile precursors and their gas-phase transformation is believed to be a significant step leading to their presence in remote regions.8 Surface waters, particularly oceans, have been identified as the major sink of PFCs.9
Pacific harbor seals (Phoca vitulina richardii) and double-crested cormorants (Phalacrocorax auritus) are year-round residents of San Francisco Bay. Because these seals and cormorants exhibit site fidelity, and forage, reproduce and nurture young in some of the more industrialized areas of the San Francisco Bay, they represent excellent indicators of Bay health and the uptake of contaminants.
Since the passage of the Marine Mammal Protection Act in 1972, harbor seal populations have dramatically increased along the California coast; the state-wide population is estimated to be 34,000.10 In contrast, seal numbers within the Bay have not changed markedly since the 1970s especially in the South Bay (estimated at approximately 500 seals).11 The possible causes for this lack of recovery are unclear and include: loss of habitat, disturbance, changes in prey availability, and contaminants.11,12
While limited data exist on the concentrations of polychlorinated biphenyls (PCBs), polybrominated diphenyl ethers (PBDEs) and pesticides in San Francisco Bay seals11–13 and cormorants,14 no data for PFCs are currently available. In addition, there is recent evidence suggesting that PFCs may cause deleterious health effects to marine mammals including suppression of the immune system, alteration of lipid metabolism, and induction of the proliferator-activated receptor-alpha (PRAR-alpha) pathway.15–19 Similar adverse outcomes have been identified for avian exposure to PFCs including reduced hatching success, reduced body weight, and increased liver weight.20–22
The goal of this project was to evaluate whether PFCs are at levels of concern in apex predators residing in a heavily urbanized estuary. Two apex predators are commonly monitored in San Francisco Bay, cormorants and seals; these species were monitored in this study to determine whether PFCs were accumulating in the Bay Area food webs. In addition, prey species of these animals were also evaluated to determine possible sources of PFC to the food chain and whether there were areas of particular concern in the estuary. A recent study of San Francisco Bay seals suggests that they are carnivorous opportunists, which consume a variety of fish including gobies (Gobiida), topsmelt (Atherinops affinis) and anchovies (Engraulis mordax), and that their diets parallel the relative abundance of fish which is a function of habitat.23 The diet of San Francisco Bay double-crested cormorant is not known; however, based on studies of cormorants in other locations, it is likely it is almost exclusively fish and will also vary according to habitat. Cormorants that reside in shallow water habitats typically consume small fish such as gobies, and sculpin; those that reside in deeper water tend to consume larger fish such as white croaker (Micropogonias furnieri).24
To assess the potential impacts of PFCs on apex predators residing in urban estuaries, 70 tissue samples were collected between 2006 and 2009 from two seal colonies and three cormorant colonies residing in the Bay and one reference seal colony. To evaluate potential sources of PFC from prey, small fish were collected from 10 sites in the Bay and one reference location.
Experimental section
Field samples
In the fall of 2009, small prey fish were sampled using beach seines from ten intertidal and subtidal sites around the margins of San Francisco Bay and one reference site in Tomales Bay (Fig. 1). Topsmelt (Atherinops affinis) were collected at eight of the sites, and Mississippi silverside (Menidia audens) were collected at six. All fish were segregated by species and measured for total length and mass. Lengths of the fish were targeted to young of the year and the length varied between 40 and 80 mm. Composite samples of seven to fifteen fish of each species were prepared in the field. The fish were wrapped in foil and transported on ice to the laboratory where they were frozen at − 20 °C. Samples were shipped on ice by overnight courier to AXYS Analytical (Sidney, BC Canada) for homogenization and analysis.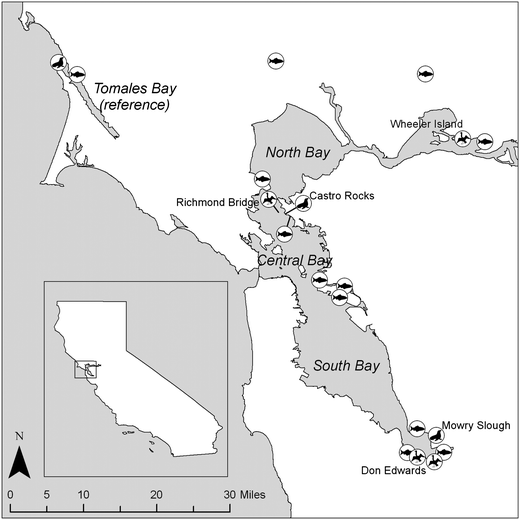 | ||
| Fig. 1 Location of seal, bird, and small fish sampling sites. | ||
Although seals move within and even out of the Bay, they tend to spend the majority of their time at the same locations.11 Samples were collected from seals residing in two of the three largest haul-out sites in San Francisco Bay, Castro Rocks and Mowry Slough.11 Haul-out sites are rocky outcroppings, tidal mudflats, and sandy shorelines that seals use to rest, molt, pup, nurse and forage. The Castro Rocks haul-out site lies in the northern portion of the Bay directly beneath the Richmond Bridge (37.9339 N, 122.4109 W); the Mowry Slough haul-out site is located in the southern portion of the Bay in the Don Edwards National Wildlife Refuge (37.493 N, 122039 W). Samples were also collected from a reference haul-out site located approximately 40 kms north of San Francisco Bay in Tomales Bay located in Point Reyes National Seashore (38.2191N, 122.9616W) (Fig. 1). Tomales Bay is an ideal reference site given the similarity of the environment of the two estuaries but the absence of any significant industrial, commercial and residential areas in the national park. All seal samples were collected under the NOAA-NMFS permit # 555-1870-00 issued to Dr James Harvey of Moss Landing Marine Laboratories, Moss Landing, California in accordance with State and Federal laws.
Thirty-four seals were captured at Castro Rocks in August of 2006, and May and June of 2007 and 2008. Another six samples of archived blood from harbor seals collected in 2004 from Mowry Slough were analyzed for PFCs. Twenty-one Tomales Bay seals were collected in May and June of 2007 and 2008. All seals were captured using tangle/seine nets25 and released to the wild after sampling.
At the time of capture, seals were weighed and measured. Blood was drawn into sterile vials using a vacutainer system (Becton Dickinson, Franklin Lakes, NJ) and stored at −4 °C for less than 8 h, centrifuged and the serum decanted into polypropylene tubes and frozen at −20 °C until analysis at AXYS Analytical. Age class was assigned based on weight and time of year: weaner (1 to 12 months), yearling (1 to 2 years), subadult (2 to 5 years), and adult (see Supplemental Information for sample sizes by age).
Cormorant eggs were collected in the Spring (March–May) in 2006 and 2009 from three locations in San Francisco Bay (Fig. 1). An appropriate reference site was not identified for cormorants. The Wheeler Island site was located in the northern portion of the Bay near the confluence of the San Joaquin and Sacramento rivers (N38.08018, W121.965). The Richmond Bridge site was located directly above the Castro Rocks seal haul-out site (N37.93684, W122.427) and the Don Edwards National Wildlife refuge was near the Mowry Slough seal site (N37.44109, W122.044; N37.4505, W122.086). Eggs were collected in accordance with State and Federal laws under the following permits: California State Migratory Bird Scientific Collection permit No. MB173904-0 issued to Dr Josh Ackerman and Federal Migratory Bird Scientific Collection permits issued by California Department of Fish and Game (CDFG) permits, SC-004741 issued to Dr Collin Eagle-Smith and SC-10107 issued to Dena Spatz.
Eggs were collected by hand (one from each nest) and aged by flotation. An effort was made to collect the freshest eggs possible. Eggs were labeled, placed into polyethylene bags, set into fiber egg cartons, and stored on ice until placement in a refrigerator at 4 °C. Samples were shipped cold by overnight courier to AXYS. At the laboratory, the eggs were weighed and composited by site, with each site having three composites consisting of seven eggs each. Eggs were stored at 4 °C until analysis.
Sample extraction and instrumental analysis
Seal serum samples (0.5 g each) were spiked with isotope-labeled surrogate standards (13C4 PFOS, 13C2 PFOA and 13C2 perfluorodecanoate (PFDA)). The samples were then mixed with a 50% aqueous solution of formic acid prior to extraction and cleanup on an SPE cartridge (Waters Oasis WAX, 60 mg and 150 mg) with methanol as the eluent. Egg homogenates (2.0 g sample size) and fish tissue (2.0 g sample size) were spiked with isotope-labeled surrogate standards (13C4 perfluorobutanoate (PFBA), 13C2 perfluorohexanoate (PFHxA), 13C2PFOA, 13C5 perfluorononanoate (PFNA), 13C2 PFDA, 13C2 perfluorododecanoate (PFDoA) and 13C4 PFOS). Samples were then agitated in a basic methanol solution and centrifuged. An aliquot of the supernatant was diluted with water and spiked with recovery standards (18C8 PFOA for egg homogenates and 13C2 2H-perfluoro-2-decenoic acid for fish tissue) prior to cleanup on an SPE cartridge with methanol as the eluent.Concentrations of PFOS, PFOA, PFBA, perfluoropentanoate (PFPeA), PFHxA, perfluoroheptanoate (PFHpA); PFNA, PFDA, perfluoroundecanoate (PFUnA), PFDoA, perfluorobutanesulfonate (PFBS), perfluorohexanesulfonate (PFHxS) and perfluorooctanesulfonamide (PFOSA) were measured using a high performance liquid chromatograph (Waters 2795, Milford MA) with a triple quadrupole mass spectrometer (Micromass Quattro Ultima, Manchester, UK) operating in the electrospray negative mode. Data were acquired using MassLynx v.4.1 software. A 15 μl sample was injected onto an Agilent Zorbax XDB reverse phase column (C18, 7.5 cm, 2.1mm id, 3.5 micron particle size column, Agilent, Santa Clara, CA). Acetonitrile/water and ammonium acetate/acetic acid were used as the mobile phases.
The following target compound transitions (parent - daughter) were monitored using Multiple Reaction Mode: perfluorobutanoate (PFBA, 213 to 169); pefluoropentanoate (PFPeA, 263 to 219); perfluorohexanoate (PFHxA, 313 to 269); perfluoroheptanoate (PFHpA, 363 to 319); PFOA (413, to 369/219); perfluorononanoate (PFNA, 463 to 419); perfluorodecanoate (PFDA, 513 to 469); perfluoroundecanoate (PFUnA, 563 to 519); perfluorododecanoate (PFDoA, 613 to 569); perfluorobutanesulfonate (PFBS, 299 to 80/99); perfluorohexanesulfonate (PFHxS, 399 to 80/99), PFOS (499 to 80/99), and perfluorooctanesulfonamide (PFOSA, 498 to 78). Results were corrected for losses during extraction and cleanup through the use of internal standards. Solvents used were of HPLC grade or better and purchased from VWR International, Mississauga, ON. Isotope-labeled standards (13C 4PFBA, 13C 2PFHxA, 13C 2PFOA, 13C 5PFNA, 13C 2PFDA, 13C 2PFDoA, 13C 4PFOS, 18C 8PFOA and 13C 2 2H-perfluoro-2-decenoic acid) were purchased from Wellington Labs and Perkin Elmer. Native standards were purchased from Sigma Aldrich and Fluka.
Target analytes for all samples included PFBA, PFPeA, PFHxA, PFHpA, PFOA, PFNA, PFDA, PFUnA, PFDoA, PFBS, PFHxS, PFOS, and PFOSA. In addition, small fish were analyzed for the following PFCs precursors: N-ethyl-perfluorooctanesulfonamide, N-ethyl-perfluorooctanesulfonamidoethanol, N-methyl-perfluorooctanesulfonamide, N-methyl-perfluorooctanesulfonamidoethanol. None of the precursors were detected. PFOSA was not detected in any seal sample and in some instances the laboratory spike for the seals exceeded the laboratory percent recovery control limits (these values were rejected). PFOSA was only detected in one bird egg sample near the MDL. As a result, the PFOSA results are not reported for seals and birds. PFOSA was detected in two small fish samples at relatively low concentrations. At the time of analysis radiolabelled PFOSA was not available; therefore this result should be viewed somewhat qualitatively.
Method detection limits (MDLs) were developed as described in the Code of Federal Regulations, Vol. 40, Part 136, Appendix B. Initial calibration occurred daily using a six-point calibration curve; a midlevel calibration standards were run after every 20th sample. All data analyses conformed to the QA/QC protocols of the laboratory and external review by SFEI.
Statistical analysis
Seal, bird, and fish data were evaluated for statistical significance using analysis of variance (ANOVA, p < 0.05) with fixed effects for site (all), time (birds), sex (seals) and age (seals). One half the MDL was used for values below the MDL. Data were tested for normality using the Kolmogorov-Smirnov test. Seal and bird data were log normal and were transformed prior to analysis with two exceptions: bird egg PFOS and PFNA data did not require log-transformation to achieve normality. An evaluation of interaction effects between variables was conducted and with the exception of PFDoA in seals, no significant interactions were identified (PFDoA exhibited an interaction between age and site). All tests were performed in Systat v. 11 (Systat Software, Inc. Chicago, IL).Results
Small fish
Three PFC compounds were detected in small fish (Table 1). PFOS was detected in 9 of the 14 composited samples in concentrations ranging from 5.7 to 80 ng g−1 ww (see Table 1 for summary and Supplemental Information for individual results). Higher mean concentrations were observed at the South Bay sites relative to Central and North Bay sites; however, this was not statistically significant. PFOSA was detected in two fish samples; one of which also contained PFDA. None of the PFC precursors were detected in small fish. No PFCs were detected in samples from the reference site.| PFHxS | PFOS | PFHpA | PFOA | PFNA | PFDA | PFUnA | PFDoA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Matrix | Year | Site | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) | FD | Mean (SD) |
| a SB- South Bay.; CB- Central Bay; NB- North Bay; Ref- reference site north of San Francisco Bay. b ND - Analyte not detected above MDL. FD - Frequency of detection. c Indicates a single value detected. No average was calculated. d Non-detect concentrations are treated as one-half the MDL for purposes of calculating mean and standard deviations. | ||||||||||||||||||
| Small fish | 2009 | SB | ND | 3/3 | 43.4 (16.1) | ND | ND | ND | ND | ND | 1/3 | 3.4 | ||||||
| CB | ND | 2/5 | 6.67 (5.79) | ND | ND | ND | ND | ND | ND | |||||||||
| NB | ND | 4/5 | 21.2 (33.0) | ND | ND | ND | ND | ND | ND | |||||||||
| Tomales Bay - Ref | ND | ND | ND | ND | ND | ND | ND | ND | ||||||||||
| Seal | 2004 | SB- Mowry | 6/6 | 64.7 (49.2) | 6/6 | 1044 (595) | 3/6 | 0.58 (0.43) | 6/6 | 7.83 (2.82) | 6/6 | 18.0 (13.7) | 6/6 | 6.60 (6.00) | 6/6 | 8.77 (5.11) | 6/6 | 6.69 (3.55) |
| 2006 2007 2008 | CB-Castro Rocks | 34/34 | 37.4 (26.4) | 34/34 | 301 (273) | 10/34 | 0.61 (0.70) | 30/34 | 2.85 (2.69) | 34/34 | 11.7 (8.4) | 34/34 | 9.66 (8.30) | 34/34 | 6.28 (4.80) | 31/34 | 4.02 (3.84) | |
| 2007 2008 | Tomales Bay - Ref | 16/21 | 2.40 (2.45) | 21/21 | 35.5 (28) | ND | 1/21 | 1.07 | 21/21 | 2.56 (7.03) | 20/21 | 1.93 (1.63) | 21/21 | 5.08 (4.51) | 14/21 | 0.71 (0.58) | ||
| Bird eggs | 2006 | SB- Don Edwards | 3/3 | 10.4 (2.6) | 3/3 | 1253 (327) | ND | 3/3 | 7.91 (2.47) | 3/3 | 13.4 (3.2) | 3/3 | 22.0 (3.6) | 3/3 | 8.91 (0.41) | 3/3 | 12.8 (7.7) | |
| CB-Richmond Bridge | 2/3 | 2.84 (0.14) | 3/3 | 280 (364) | ND | ND | 1/3 | 5.19 | 1/3 | *7.97 | 3/3 | 4.84 (1.40) | 3/3 | 5.96 (2.05) | ||||
| NB-Wheeler Island | ND | 3/3 | 104 (30) | ND | 1/3 | 2.70 | 3/3 | 7.04 (1.35) | 3/3 | 8.4 (2.7) | 3/3 | 7.15 (2.56) | 3/3 | 6.09 (2.64) | ||||
| 2009 | SB- Don Edwards | 3/3 | 25.2 (13.2) | 3/3 | 1244 (472) | ND | 3/3 | 24.3 (3.9) | 3/3 | 30.5 (7.8) | 3/3 | 22.9 (4.7) | 3/3 | 7.84 (1.40) | 3/3 | 7.6 (2.7) | ||
| CB-Richmond Bridge | 2/3 | 4.44 (1.76) | 3/3 | 123 (32) | ND | 1/3 | 5.92 | 3/3 | 3.81 (0.87) | 3/3 | 8.9 (4.6) | 3/3 | 7.17 (3.55) | 3/3 | 9.6 (5.2) | |||
| NB-Wheeler Island | ND | 3/3 | 84 (11) | ND | ND | 3/3 | 5.90 (1.78) | 3/3 | 9.52 (2.60) | 3/3 | 4.00 (1.02) | 3/3 | 4.05 (0.99) | |||||
Seals
Eight PFCs were detected in seal serum (Table 1). PFOS, PFNA, and PFUnA were detected in all samples. Average concentrations of PFOS were generally an order of magnitude higher than those of other PFCs. The second highest observed concentrations were of PFHxS with a mean concentration of 65 ng mL−1 from Mowry Slough (South Bay) and 37 ng mL−1 from Castro Rocks (Central Bay). Concentrations of the remaining compounds were less than 20 ng mL−1. Concentrations of PFCs were greater in the Mowry Slough seals than the Castro Rocks seals with one exception (PFDA). Seals from San Francisco Bay had higher concentrations than seals from the reference site. The differences among the sites (South Bay vs. Central Bay vs. Tomales Bay) were statistically significant for all compounds (p = 0.001) except PFHpA which was significant (p = 0.01) and PFUnA which was not significant.Cormorants
Seven compounds were detected in cormorant eggs (Table 1). Average concentrations of PFOS were generally an order of magnitude higher than those of the other PFCs. Longer chain PFCAs such as PFNA, PFDoA, and PFUnA were the next most prevalent compounds, average concentrations ranged from 3.8 to 30 ng g−1. PFHxS was largely only detected in the South Bay and average concentrations increased from 10.4 in 2006 to 25.2 in 2009. Concentrations of PFHxS across all sites for the two sampling events were statistically significant (p = 0.05). As with seals, concentrations of PFCs in eggs were always greater in the South Bay (p = 0.001) and PFOA was always detected in the South Bay site and sporadically detected at the remaining two sites. Concentrations of PFOS remained virtually identical between the two sampling events.Discussion
Small fish
Consistent with previous studies,3,26,27 PFOS was the main PFC detected in small fish in this study. The average concentration of PFOS observed in San Francisco Bay is in the range of values reported for prey fish elsewhere (Table 2). Interestingly, an order of magnitude higher concentration was observed in Lake Ontario sculpin, a benthic-dwelling fish.27 In general, sulfonates tend to have a higher bioaccumulation potential in fish than the carboxylates and PFCs with fewer than eight carbons in the perfluoroalkyl chain tend not to bioaccumulate.28,29 These patterns were confirmed in this study.| Species | N | Mean (std dev) | Location | Collection Date | Reference |
|---|---|---|---|---|---|
| Topsmelt (Atherinops affinis)/Silversides (Medina audens) | 13 | 20.7(25.0) | San Francisco Bay | 2009 | This study |
| Topsmelt (Atherinops affinis) | 1 | ND | Reference | 2009 | This study |
| Herring | 10 | 2.3 | Baltic Sea | 1989 | 51 |
| Slimy sculpin (Cottus cognatus) | 5 | 450 (98) | Lake Ontario | 2002 | 27 |
| Alewife (Alosa pseudoharengus) | 6 | 46 (15) | Lake Ontario | 2002 | 27 |
| Smelt (Osmerus mordax) | 6 | 110 (55) | Lake Ontario | 2002 | 27 |
| Round gobies (Neogobius melanostomus) | 12 | 10.6 (5.3) | Rivers near Great Lakes | 1998–1999 | 26 |
| Pinfish (Lagodon rhomboids) | 10 | 4.8 (4.5) | Florida US | 2004 | 37 |
| Pigfish (Orthopristis chrysoptera) | 10 | 3.1 (2.5) | Florida US | 2004 | 37 |
Higher PFOS concentrations were observed in Mississippi silversides relative to topsmelt (33.1 vs. 10.1 ng g−1, see Supplemental information); however, this difference was not statistically significant. Silversides inhabit the shallow tidal areas while topsmelt frequent areas of open water. Higher concentrations of PFOS also were observed in Lake Ontario benthic-dwelling fish (e.g., sculpin) than in pelagic fish (alewife and smelt). This difference was attributed to diet, specifically benthic-dwelling shrimp which have relatively high concentrations given their trophic status.27
PFOSA, which was detected in 2 of the 14 composites at concentrations of 4.3 and 9.6 ng g−1, also has been reported in prey fish collected in the Great Lakes and Europe.3,26,27 Fish are able to metabolize precursors such as N-ethyl perfluorooctanesulfonamide to PFOS, demonstrating that the mechanism to degrade PFOSA and other precursors exist in fish4,27 and suggesting that some of the PFOS detected in fish may be due to the metabolism of precursors.
Seals
PFOS was the main PFC detected (Table 1), corroborating previous data for seals,30–35 harbor porpoises (Phocoena phoceoena36), and bottlenose dolphins (Tursiops truncatus,37,38). PFOS concentrations observed in serum in this study were higher than previously reported values for seals including ringed, gray, harbor, elephant, and Baikal (Table 3). The average PFOS concentrations for San Francisco Bay seals (412 ng mL−1) are slightly higher than concentrations observed in blood of four stranded harbor seals collected off the coast of northern Germany (349 ng g−1 ww,39). However, stranded animals are less likely to be representative of the overall population as a result of emaciation and compromised immune systems.| Species | Sample | Units | n | Mean (std dev) | Range | Location | Date | Reference |
|---|---|---|---|---|---|---|---|---|
| Harbor seals (Phoca viulina) | Serum | ng/mL | 40 | 412 (425) | 1960–16.5 | San Francisco Bay, CA | 2004–2008 | This study |
| Serum | ng/mL | 21 | 35.5 (28) | 134–13.5 | Tomales Bay, CA | 2007–2008 | This study | |
| Blood | ng/g | 4 | 349 (370) | 48–887 | German Bight | 2007 | 39 | |
| Ringed seals (Phoca hispida) | plasma | ng/ml | 18 | 110 | 16–230 | Baltic Sea | 1990s | 2 |
| plasma | ng/mL | 18 | 9 | 5–14 | Norwegian Arctic | 1990s | 2 | |
| Blood | ng/g | 5 | 2.5–8.6 | Canadian Arctic | 2004 | 33 | ||
| Gray seals | plasma | ng/mL | 26 | 37 | 14–76 | Baltic Sea | 1990s | 2 |
| plasma | ng/ml | 12 | 28 | 11–49 | Canadian Arctic | 1990s | 2 | |
| Baikal seals (Pusa sibirica) (male) | Serum | ng/g | 19 | 5.8 (4.9) | 1.4–17 | Lake Baikal | 2005 | 15 |
| (female) | Serum | ng/g | 24 | 4.5(3.6) | 1.3–1.4 | Lake Baikal | 2005 | 15 |
| Elephant seals (Mirounga leonine) | Blood | ng/mL | 59 | 0.53 | <0.08–3.52 | Antarctic | 2004–2005 | 67 |
| Bottlenose Dolphins (Tursiops truncates) | plasma | ng/g | 13 | 658 | 194–1715 | Sarasota Bay, FL | 2003 | 38 |
| plasma | ng/g | 42 | 462 | 69–2019 | Indian River Lagoon, FL | 2003 | 38 | |
| plasma | ng/g | 47 | 1171 | 472–3073 | Charleston, SC | 2003 | 38 | |
| plasma | ng/g | 5 | 646 | 232–1240 | Delaware Bay | 2003 | 38 | |
| plasma | ng/g | 2 | 49 | 46–52 | Bermuda | 2003 | 38 |
The highest reported PFOS concentrations in marine mammals have been observed in polar bears and dolphins.3,27,37 Average concentrations of PFOS in blood from dolphins collected in Charleston Harbor were 1,171 ng g−1 ww38 and are similar to the values observed for the South Bay seals. Because polar bears eat seals and the concentrations of PFOS in arctic seals is substantially lower, these results suggest that marine mammals residing in urban estuaries such as Charleston Harbor, Delaware Bay, and Guanabara Bay (Brazil) have a substantially higher PFOS burden than their counterparts in remote locations.3,38,40,41
Information on the toxicity of PFOS to seals is limited. Thresholds for effects from PFOS exposure for other mammals are several orders of magnitude higher than the concentrations observed in this study (e.g., no adverse effect levels of 82.6 ug/g in serum for male monkeys,42 and benchmark dose for neonatal survival for rats of 25 ug/g in serum18). However, a number of researchers have recently reported adverse effects at concentrations comparable to those observed in San Francisco Bay seals. For example, reduced birth weights and head circumferences in humans were observed at a median population concentration of PFOS of 5 ng mL−1 cord blood.43 Suppression of the immune system in seals, rats, and sea otters has also been observed.15,17,44,45 Guruge et al.44 observed a significant increase in emaciation and mortality in a study on the effects of PFOS on rat resistance to influenza. Rats that had 670 ng ml−1 of PFOS in their plasma had a 17 percent survival as compared to 46 percent survival for the control (2.1 ng ml−1). In a study of California sea otters, Kannan et al.17 reported a significant correlation between the incidence of disease and PFOS/PFOA concentrations in livers (PFOS concentrations ranged 1 to 884 ng g−1 ww).
PFHxS was consistently detected at high concentrations in seals from the South Bay. This is a somewhat anomalous finding as PFNA is usually the second most frequently detected PFC in pinipeds.15,31,32,39,46 It was also detected in relatively high concentrations in bird eggs. PFHxS has been identified as an ingredient in aqueous film-forming foams for fire suppression (AFFFs)47 and carpet treatment applications.48 The prevalence of PFHxS in this study may reflect local sources such as military, aviation, and wastewater treatment facilities.
The PFCs with fewer than six carbons were not detected in any sample which is consistent with previous studies suggesting their low bioaccumulation potential.28,29,32 The detection of PFCAs was correlated strongly with elevated concentrations of PFOS (p = 0.001). Average PFOA concentrations in the Mowry Slough and Castro Rocks seals were low, 7.8 ng mL−1 and 2.8 ng mL−1, respectively. With one exception, PFOA was not detected in the seals from the reference site. These results are consistent with the literature30–32,35 and the hypothesis that PFOA has a low bioaccumulation potential.28,29
No statistically significant difference in PFOS was observed among males and females. The average PFOS concentration detected in male seals was 299 ng mL−1 (n = 29); the average concentration for females was 268 ng mL−1 (n = 32, see Supplemental Information). No sex differences have been reported by other researchers3,35,36,41,46 except for Ishibashi et al.15 who observed statistically higher concentrations in male serum versus female serum for Lake Baikal seals.
PFC concentrations did not increase with age. These results are consistent with prior seal studies32,34,35,46 and the relatively short-half lives of PFCs in marine mammals. The estimated half-life of PFOS in bottlenose dolphins is 21 weeks.3 Although PFCs do not appear to increase with age, some researchers have observed significantly elevated concentrations in recently weaned seals.15,46 In this study, PFOS concentrations in weaned pups were statistically higher than yearlings and subadults (p = 0.001). The higher concentration may be due to lactational or in utero transfers.15 PFCs have been detected in dolphin milk3 and elevated fetal PFC concentrations relative to the mothers have been observed in seals, dolphins and melon-headed whales.31,40,46,49 These results suggest that both mechanisms are plausible.
Cormorant eggs
PFOS was the primary PFC detected in cormorant eggs (Table 1). This is consistent with previous bird data.50–53 Average PFOS concentrations in the South Bay are 1,249 ng g−1 ww and were between 2 and 16 times higher than values reported elsewhere (Table 4). Average PFOS concentrations in Central Bay/Richmond Bridge and North Bay/Wheeler Island eggs were lower (202 and 113 ng g−1 ww, respectively) and are within the range reported by other researchers.51–55| Species | n | Mean (std dev) | Range | Location | Collection Date | Reference |
|---|---|---|---|---|---|---|
| Double-crested cormorant (Phalacrocorax auritus) | 6 | 1249 | 833–1760 | South Bay | 2006/2009 | This study |
| 6 | 202 | 63–700 | Central Bay | 2006/2009 | This study | |
| 6 | 94 | 73.8–135 | Delta | 2006/2009 | This study | |
| 4 | 157 | 21–220 | Manitoba, Canada | 1995 | 41 | |
| 4 | 210 | 130–210 | Lake Winnipeg, Canada | 1990s | 2 | |
| Common Guillemot (Uria aalge) | 8 | 325 | 243–432 | Baltic Sea | 1989 | 51 |
| 9 | 614 | 551–669 | Baltic Sea | 2003 | 57 | |
| 10 | 400 | 200–760 | Baltic Sea | 2003 | 52 | |
| 10 | 16 | 5.2–22 | Iceland | 2002 | 52 | |
| Ring-billed gull (Larus delawarensis) | 3 | 67 | 30–126 | Lake Huron, Michigan | 1995 | 41 |
| Herring gull (Larus argentatus) | 13 | 586 (62) | Toronto Harbor, Canada | 2007 | 55 | |
| Herring gull (Larus argentatus) | 13 | 148(22) | Granite Island, Lake Superior | 2007 | 55 | |
| Ivory gull (Pagophila eburnea) | 10 | 79.2 | 24.2–113 | Norwegian Arctic | 2007 | 54 |
| Night heron (Nycticorax nycticorax) | 9 | 115 | 22.6–343 | Hong Kong | 2006 | 53 |
Adverse effects associated with avian exposure to PFOS include: reduced body weight; an increase in liver weight; an increase in incidence of small testes for northern bobwhite quail (Colinus virginianus) and mallard (Anas playrhynchos);21 and reduced hatching and pipping success in chickens (Gallus domesticus).20,22 Based on effects on offspring survival of northern bobwhite quails, Newsted et al.21 developed a predicted no effect concentration (PNEC) of 1 μg mL−1 for eggs. Effects to hatching and pipping success ranged from 0.1 μg PFOS/g egg (lowest observed adverse effect level20) to 100 μg/g.22 An additional concern is that PFCs appear to enhance the toxicity of dioxin and dioxin-related compounds in wild cormorants.56 While there is uncertainty regarding the relative sensitivity of cormorants to PFCs, Newsted et al.21 suggested that the inter-species differences may not be significant.
Conclusions
Seals and cormorant eggs collected from the South Bay contained higher average concentrations of PFOS than those reported for other sites. The elevated concentrations of PFOS in the South Bay and the absence of a decrease following the curtailment in production in 2001 in the US suggest continued sources, particularly given that PFOS has relatively short half-lives in dolphins (21 weeks,3) and birds (two to three weeks,21). The spatial trend is likely due to several factors including: the limited circulation in the southern portion of the Bay; the greater influence of stormwater runoff, wastewater effluent, and other potential sources; and the different type of habitats/prey species present in each area (tidal mudflats and shallow water versus rocky outcrops in relatively deep water).The absence of a decline in PFOS concentrations in bird eggs over time is in contrast to several recent reports documenting the decline of PFCs in biota elsewhere including seals in the Arctic,35 sea otters along the California coast,17 birds in the Baltic Sea,57 and humans in the US.58 Some of these declines have been dramatic; for example, a 60 percent decrease in PFOS concentrations was observed in human blood donors from six centers across the US over the period 2000 to 2006.58 However, decreases in PFOS in biota are by no means universal30,49 and the absence of a decline has been attributed to on-going sources.
Sources of PFCs to the Bay remain to be elucidated. A number of land uses and pathways that have been historically associated with PFCs are located along the Bay shoreline including airports, military facilities, landfills, wastewater treatment plants, oil refineries, and stormwater discharges.3,47,59,60 PFCs are also present in consumer products including treated textiles, carpets, and food-packaging materials, and diffuse releases from homes and commercial facilities represent yet another potential source.3,61,62 In addition, although perfluorooctane sulfonyl fluoride (POSF) based compounds were phased out in 200163 and the USEPA restricted the use of perfluoroalkyl sulfonates in the 2002 Significant New Use Ruling,64 a number of uses were exempted such as the use of perfluoroalkyl sulfonates as an anti-erosion additive used in aviation hydraulic fluid, a component of photo-resist in the semi-conductor/electronics industry, and a coating for surface tension applications in analog and digital imaging films, plates, and paper. It is not known whether these uses are significant in the Bay Area. Finally, a previous study of San Francisco Bay sediments identified PFOS and PFC precursors such as 2-(N-ethylperfluorooctane sulfonamide) acetic acid and 2-(-N-methylperfluorooctane sulfonamide) acetic acid in concentrations greater than or equal to PFOS65 suggesting that the sediments may also be a potential contribution.
The concentrations of PFOS observed in cormorant eggs and seals are within the range where adverse effects have been observed. It is not known whether these will translate into population effects. However, the seal colony at Mowry Slough has not shown a significant increase since the 1970s;11 this is in marked contrast to similar sites in Point Reyes National Seashore where there has been an increase in population.66 The reasons for this lack of increase are unknown; it will be important to elucidate if contaminants such as PFCs are a factor.
Acknowledgements
The authors would like to thank the Regional Monitoring Program for Water Quality in San Francisco Estuary, the Valentine Family Foundation and the John H. Prescott Marine Mammal Rescue Assistance Grant Program for funding; Rachel Allen and Jay Davis for technical review and assistance; Aroon Melwani for statistical analysis; Josh Ackerman and Collin Eagles-Smith of USGS-WREC for collection of bird eggs; The Marine Mammal Center and Moss Landing Marine Labs for the collection of seal samples; Ben Greenfield and Kat Ridolfi of SFEI and Darrell Slotton of UC-Davis for collection of small fish; and Richard Grace and Candice Navaroli of AXYS Analytical for PFC analyses.References
- E. Kissa, Fluorinated Surfactants and Repellents. 2nd ed, 2001, New York: Marcel Dekker Search PubMed.
- J. P. Giesy and K. Kannan, Global distribution of perfluorooctane sulfonate in wildlife, Environ. Sci. Technol., 2001, 35(7), 1339–1342 CrossRef CAS.
- M. Houde, et al., Biological monitoring of polyfluoroalkyl substances: A review, Environ. Sci. Technol., 2006, 40(11), 3463–3473 CrossRef CAS.
- G. T. Tomy, et al., Biotransformation of N-ethyl perfluorooctanesulfonamide by rainbow trout (Onchorhynchus mykiss) liver microsomes, Environ. Sci. Technol., 2004, 38(3), 758–762 CrossRef CAS.
- J. C. D'Eon, et al., Atmospheric chemistry of N-methyl perfluorobutane sulfonamidoethanol, C4F9SO2N(CH3)CH2CH2OH: Kinetics and mechanism of reaction with OH, Environ. Sci. Technol., 2006, 40(6), 1862–1868 CrossRef CAS.
- M. J. A. Dinglasan, et al., Fluorotelomer alcohol biodegradation yields poly- and perfluorinated acids, Environ. Sci. Technol., 2004, 38(10), 2857–2864 CrossRef CAS.
- D. A. Ellis, et al., Atmospheric lifetime of fluorotelomer alcohols, Environ. Sci. Technol., 2003, 37(17), 3816–3820 CrossRef CAS.
- J. W. Martin, et al., Atmospheric Chemistry of Perfluoroalkanesulfonamides: Kinetic and Product Studies of the OH Radical and Cl Atom Initiated Oxidation of N-Ethyl Perfluorobutanesulfonamide, Environ. Sci. Technol., 2006, 40(3), 864–872 CrossRef CAS.
- K. Prevedouros, et al., Sources, fate and transport of perfluorocarboxylates, Environ. Sci. Technol., 2006, 40(1), 32–44 CrossRef CAS.
- J. V. Carretta, et al., U.S. Pacific Marine Mammal Stock Assessments: 2007, 2007, U.S. Department of Commerce, National Oceanic and Atmospheric Administration: La Jolla, CA. p. 316. Search PubMed.
- E. K. Grigg, et al., Harbor seal, Phoca vitulina richardii, population trends in the San Francisco Bay estuary, 1970–2002, California Fish and Game, 2004, 90(2), 51–70 Search PubMed.
- J. C. C. Neale, et al., Contaminant loads and hematological correlates in the harbor seal (Phoca vitulina) of San Francisco Bay, California, J. Toxicol. Environ. Health, Part A, 2005, 68(8), 617–633 CrossRef CAS.
- J. W. She, et al., PBDEs in the San Francisco Bay Area: measurements in harbor seal blubber and human breast adipose tissue, Chemosphere, 2002, 46(5), 697–707 CrossRef CAS.
- J. A. Davis, et al., Monitoring Pollutant Concentrations in Eggs of Double-crested Cormorants from San Francisco Bay in 2002 and 2004: A Regional Monitoring Program Pilot Study, 2007, San Francisco Estuary Institute: Oakland, CA Search PubMed.
- H. Ishibashi, et al., Contamination and effects of perfluorochemicals in Baikal Seal (Pusa sibirica). 1. Residue level, tissue distribution, and temporal trend, Environ. Sci. Technol., 2008, 42(7), 2295–2301 CrossRef CAS.
- H. Ishibashi, et al., Contamination and effects of perfluorochemicals in Baikal Seal (Pusa sibirica). 2. Molecular characterization, expression level, and transcriptional activation of peroxisome proliferator-activated receptor alpha, Environ. Sci. Technol., 2008, 42(7), 2302–2308 CrossRef CAS.
- K. Kannan, E. Perrotta and N. J. Thomas, Association between perfluorinated compounds and pathological conditions in southern sea otters, Environ. Sci. Technol., 2006, 40(16), 4943–4948 CrossRef CAS.
- C. Lau, et al., Perfluoroalkyl acids: A review of monitoring and toxicological findings, Toxicol. Sci., 2007, 99(2), 366–394 CrossRef CAS.
- D. J. Greig, et al., Geography and stage of development affect persistent organic pollutants in stranded and wild-caught harbor seal pups from central California, Sci. Total Environ., 2011, 409(18), 3537–3547 CrossRef CAS.
- E. D. Molina, et al., Effects of air cell injection of perfluorooctane sulfonate before incubation on development of the white leghorn chicken (Gallus domesticus) embryo, Environ. Toxicol. Chem., 2006, 25(1), 227–232 CrossRef CAS.
- J. L. Newsted, et al., Avian toxicity reference values for perfluorooctane sulfonate, Environ. Sci. Technol., 2005, 39(23), 9357–9362 CrossRef CAS.
- J. M. O'Brien, et al., Perfluorooctane sulfonate (PFOS) toxicity in domestic chicken (Gallus gallus domesticus) embryos in the absence of effects on peroxisome proliferator activated receptor alpha (PPAR alpha)-regulated genes, Comp. Biochem. Physiol., Part C: Toxicol. Pharmacol., 2009, 149(4), 524–530 CrossRef.
- C. Gibble., Food habits of Harbor Seals (Phoca Vitulina Richardii) in San Francisco Bay, in Department of Marine Sciences, 2011, San Jose State University: San Jose. p. 51 Search PubMed.
- J. H. Johnson, et al., A Comparative Analysis of Double-Crested Cormorant Diets from Stomachs and Pellets from Two Lake Ontario Colonies, Journal of Freshwater Ecology, 2010, 25(4), 669–672 CrossRef.
- S. J. Jeffries, R. F. Brown and J. T. Harvey, Techniques for capturing, handling and marking harbour seals, Aquatic Mammals, 1993, 19(1), 21–25 Search PubMed.
- K. Kannan, et al., Perfluorinated compounds in aquatic organisms at various trophic levels in a Great Lakes food chain, Arch. Environ. Contam. Toxicol., 2005, 48(4), 559–566 CrossRef CAS.
- J. W. Martin, et al., Perfluoroalkyl contaminants in a food web from lake Ontario, Environ. Sci. Technol., 2004, 38(20), 5379–5385 CrossRef CAS.
- J. W. Martin, et al., Dietary accumulation of perfluorinated acids in juvenile rainbow trout (Oncorhynchus mykiss), Environmental Toxicology And Chemistry, 2003, 22(1), 189–195 CAS.
- J. W. Martin, et al., Bioconcentration and tissue distribution of perfluorinated acids in rainbow trout (Oncorhynchus mykiss), Environmental Toxicology and Chemistry, 2003, 22(1), 196–204 CAS.
- R. Bossi, F. F. Riget and R. Dietz, Temporal and spatial trends of perfluorinated compounds in ringed seal (Phoca hispida) from Greenland, Environ. Sci. Technol., 2005, 39(19), 7416–7422 CrossRef CAS.
- K. I. Van de Vijver, et al., Tissue distribution of perfluorinated chemicals in harbor seals (Phoca vitulina) from the Dutch Wadden Sea, Environ. Sci. Technol., 2005, 39(18), 6978–6984 CrossRef CAS.
- C. M. Butt, et al., Spatial trends of perfluoroalkyl compounds in ringed seals (Phoca hispida) from the Canadian Arctic, Environ. Toxicol. Chem., 2008, 27(3), 542–553 CrossRef CAS.
- C. R. Powley, et al., Polyfluorinated chemicals in a spatially and temporally integrated food web in the Western Arctic, Chemosphere, 2008, 70(4), 664–672 CrossRef CAS.
- G. T. Tomy, et al., Trophodynamics of Some PFCs and BFRs in a Western Canadian Arctic Marine Food Web, Environ. Sci. Technol., 2009, 43(11), 4076–4081 CrossRef CAS.
- C. M. Butt, et al., Rapid response of arctic ringed seals to changes in perfluoroalkyl production, Environ. Sci. Technol., 2007, 41(1), 42–49 CrossRef CAS.
- K. I. Van de Vijver, et al., Occurrence of perfluorooctane sulfonate and other perfluorinated alkylated substances in harbor porpoises from the Black Sea, Environ. Sci. Technol., 2007, 41(1), 315–320 CrossRef CAS.
- M. Houde, et al., Biomagnification of perfluoroalkyl compounds in the bottlenose dolphin (Tursiops truncatus) food web, Environ. Sci. Technol., 2006, 40(13), 4138–4144 CrossRef CAS.
- M. Houde, et al., Polyfluoroalkyl Compounds in Free-Ranging Bottlenose Dolphins (Tursiops truncatus) from the Gulf of Mexico and the Atlantic Ocean, Environ. Sci. Technol., 2005, 39(17), 6591–6598 CrossRef CAS.
- L. Ahrens, U. Siebert and R. Ebinghaus, Total body burden and tissue distribution of polyfluorinated compounds in harbor seals (Phoca vitulina) from the German Bight, Mar. Pollut. Bull., 2009, 58(4), 520–525 CrossRef CAS.
- P. R. Dorneles, et al., High accumulation of perfluorooctane sulfonate (PFOS) in marine tucuxi dolphins (Sotalia guianensis) from the Brazilian coast, Environ. Sci. Technol., 2008, 42(14), 5368–5373 CrossRef CAS.
- K. Kannan, et al., Accumulation of perfluorooctane sulfonate in marine mammals, Environ. Sci. Technol., 2001, 35(8), 1593–1598 CrossRef CAS.
- A. M. Seacat, et al., Subchronic toxicity studies on perfluorooctanesulfonate potassium salt in cynomolgus monkeys, Toxicol. Sci., 2002, 68(1), 249–264 CrossRef CAS.
- B. J. Apelberg, et al., Cord serum concentrations of perfluorooctane sulfonate (PFOS) and perfluorooctanoate (PFOA) in relation to weight and size at birth, Environ. Health Perspect., 2007, 115(11), 1670–1676 CrossRef CAS.
- K. S. Guruge, et al., Effect of perfluorooctane sulfonate (PFOS) on influenza A virus-induced mortality in female B6C3F1 mice, J. Toxicol. Sci., 2009, 34(6), 687–691 CrossRef CAS.
- M. M. Peden-Adams, et al., Suppression of humoral immunity following exposure to the perfluorinated insecticide sulfluramid, J. Toxicol. Environ. Health, Part A, 2007, 70(13–14), 1130–1141 CrossRef CAS.
- S. Shaw, et al., Specific accumulation of perfluorochemicals in harbor seals (Phoca vitulina concolor) from the northwest Atlantic, Chemosphere, 2009, 74(8), 1037–1043 CrossRef CAS.
- C. A. Moody, et al., Monitoring perfluorinated surfactants in biota and surface water samples following an accidental release of fire-fighting foam into Etohicoke Creek, Environ. Sci. Technol., 2002, 36(4), 545–551 CrossRef CAS.
- G. W. Olsen, et al., Historical comparison of perfluorooctanesulfonate, perfluorooctanoate, and other fluorochemicals in human blood, Environ. Health Perspect., 2005, 113(5), 539–545 CrossRef CAS.
- K. Hart, et al., Time trends and transplacental transfer of perfluorinated compounds in melon-headed whales stranded along the Japanese coast in 1982, 2001/2002, and 2006, Environ. Sci. Technol., 2008, 42(19), 7132–7137 CrossRef CAS.
- W. A. Gebbink, C. E. Hebert and R. J. Letcher, Perfluorinated Carboxylates and Sulfonates and Precursor Compounds in Herring Gull Eggs from Colonies Spanning the Laurentian Great Lakes of North America, Environ. Sci. Technol., 2009, 43(19), 7443–7449 CrossRef CAS.
- K. E. Holmstrom and U. Berger, Tissue distribution of perfluorinated surfactants in common guillemot (Uria aalge) from the Baltic Sea, Environ. Sci. Technol., 2008, 42(16), 5879–5884 CrossRef.
- K. Lofstrand, et al., Spatial trends of polyfluorinated compounds in guillemot (Uria aalge) eggs from North-Western Europe, Chemosphere, 2008, 72(10), 1475–1480 CrossRef.
- Y. Wang, et al., Perfluorooctane Sulfonate and Other Fluorochemicals in Waterbird Eggs from South China, Environ. Sci. Technol., 2008, 42(21), 8146–8151 CrossRef CAS.
- C. Miljeteig, et al., High Levels of Contaminants in Ivory Gull Pagophila eburnea Eggs from the Russian and Norwegian Arctic, Environ. Sci. Technol., 2009, 43(14), 5521–5528 CrossRef CAS.
- W. A. Gebbink and R. J. Letcher, Linear and Branched Perfluorooctane Sulfonate Isomer Patterns in Herring Gull Eggs from Colonial Sites Across the Laurentian Great Lakes, Environ. Sci. Technol., 2010, 44(10), 3739–3745 CrossRef CAS.
- K. Nakayama, et al., Potential Effects of Perfluorinated Compounds in Common Cormorants from Lake Biwa, Japan: An Implication from the Hepatic Gene Expression Profiles by Microarray, Environ. Toxicol. Chem., 2008, 27(11), 2378–2386 CrossRef CAS.
- K. E. Holmstrom, U. Jarnberg and A. Bignert, Temporal trends of PFOS and PFOA in guillemot eggs from the Baltic Sea, 1968–2003, Environ. Sci. Technol., 2005, 39(1), 80–84 CrossRef.
- G. W. Olsen, et al., Decline in perfluorooctanesulfonate and other polyfluoroalkyl chemicals in American Red Cross adult blood donors, 2000–2006, Environ. Sci. Technol., 2008, 42(13), 4989–4995 CrossRef CAS.
- C. A. Moody, et al., Occurrence and persistence of perfluorooctanesulfonate and other perfluorinated surfactants in groundwater at a fire-training area at Wurtsmith Air Force Base, Michigan, USA, J. Environ. Monit., 2003, 5(2), 341–345 RSC.
- S. Taniyasu, et al., A Survey of Perfluorooctane Sulfonate and Related Perfluorinated Organic Compounds in Water, Fish, Birds, and Humans from Japan, Environ. Sci. Technol., 2003, 37(12), 2634–2639 CrossRef CAS.
- T. H. Begley, et al., Perfluorochemicals: Potential sources of and migration from food packaging, Food Addit. Contam., 2005, 22(10), 1023–1031 CrossRef CAS.
- J. C. D'Eon and S. A. Mabury, Production of perfluorinated carboxylic acids (PFCAs) from the biotransformation of polyfluoroalkyl phosphate surfictants (PAPS): Exploring routes of human contamination, Environ. Sci. Technol., 2007, 41(13), 4799–4805 CrossRef CAS.
- . 3M, Letter from William Weppner of 3M to Charles Auer of USEPA, 2000, Federal Register Search PubMed.
- U.S. Environmental Protection Agency, Perfluoroalkyl Sulfonates Significant New Use Rule. Final Rule, Federal Register, 2002, 67(236), 72854–72867 Search PubMed.
- C. P. Higgins, et al., Quantitative Determination of Perfluorochemicals in Sediments and Domestic Sludge, Environ. Sci. Technol., 2005, 39(11), 3946–3956 CrossRef CAS.
- W. J. Sydeman and S. G. Allen, Pinniped population dynamics in central California: Correlations with sea surface temperature and upwelling indices, Mar. Mamm. Sci., 1999, 15(2), 446–461 CrossRef.
- L. Tao, et al., Perfluorooctanesulfonate and related fluorochemicals in albatrosses, elephant seals, penguins, and Polar Skuas from the Southern Ocean, Environ. Sci. Technol., 2006, 40(24), 7642–7648 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c1em10609k |
| This journal is © The Royal Society of Chemistry 2012 |