
Quantum chemical studies of dioxygen activation by mononuclear non-heme iron enzymes with the 2-His-1-carboxylate facial triad
Abstract
Density functional theory with the B3LYP hybrid functional has been used to study the mechanisms for dioxygen
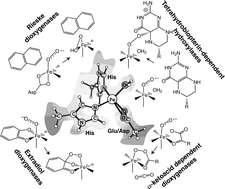
Density functional theory with the B3LYP hybrid functional has been used to study the mechanisms for dioxygen
A. Bassan, T. Borowski and P. E. M. Siegbahn, Dalton Trans., 2004, 3153 DOI: 10.1039/B408340G
To request permission to reproduce material from this article, please go to the Copyright Clearance Center request page.
If you are an author contributing to an RSC publication, you do not need to request permission provided correct acknowledgement is given.
If you are the author of this article, you do not need to request permission to reproduce figures and diagrams provided correct acknowledgement is given. If you want to reproduce the whole article in a third-party publication (excluding your thesis/dissertation for which permission is not required) please go to the Copyright Clearance Center request page.
Read more about how to correctly acknowledge RSC content.
 Please wait while we load your content...
Please wait while we load your content...

